

Immunology is the science that deals with the immune system. The immune system is a physiological system and a collection of cells and molecules which developed in the process of evolution of man, enabling him to respond to foreign antigens and tolerate his own antigens. In order to protect an individual from self-recognition for an immunological response to own antigens and an exposure to microbial infection or even an airborne pollen, the immune system evolves a set of highly complex regulatory processes, which involves a set of mechanisms to monitor the cells by immune surveillance to detect neoplastic changes in the host and to maintain the integrity of the body as a whole. Immunological response to an antigen derived from infectious and noninfectious agents is the result of a coordinated reaction of the cells and molecules of the immune system.
The progress of its growth from conceptual to real state traditionally relies on observations and the queries: what, how and why, that man puts to himself and then tries to solve. Each solution gives rise to several new queries that originate from curiosity of man and testing of the facts about nature which help in proliferation and expansion of the science. The background of this phenomenon provides a long and gradual evolution of ideas to such a system of unresponsiveness to self antigens and responsiveness to non-self antigens, that respectively give rise to tolerance and immunity with special reference to the specificity, diversity and memory (Fig. 1.1) which is of immense value for understanding the subject of immunology and teaching the speciality to the students of biomedical science.
There are several aspects of the immune phenomenon. Some knowledge of one aspect for understanding another is essential in immunology such as the reference of antibody to define antigen and the knowledge of immunity for tolerance, and also to appreciate that autoimmune disease is a mirror image of tolerance. It is also essential to know the details of lymphoid tissues for immunodeficiencies and the reference of T cell for differentiation of B cell into plasma cell in cognate interaction or linked immune recognition.
In immune recognition, the responsiveness of the lymphocytes to non-self antigens is a unique phenomenon. However, the random creation of cell surface receptors by gene rearrangement prior to antigen contact for immune recognition or an immune response and the million-fold range in binding affinities for a particular type of recognition, make the immune system intrinsically difficult to regulate.
When the regulatory mechanism of deletion of anti-self lymphocytes fails, responsiveness to self-antigen may occur and give rise to auto-immunity or auto immune disease of immunity. When the immunoregulatory mechanism is perturbed and there is a state of impaired immunity e.g., immunodeficiency, as a result of primary defect in one or more components of the immune system or secondary to certain extraneous factors (such as infection and malnutrition), the resistance to infection or immune capabilities of the individual to antigens derived from infectious agents declines and thereby the susceptibility of the host for infection increases.


Similarly, when the immunoregulatory mechanism goes wrong and there is a state of inappropriate immunity e.g., hypersensitivity, the features of altered deleterious reactivity of exaggerated immune response to one or more non-self antigen may appear that may in turn harm the host and give rise to allergy.
Immunology is one of those subjects that has made remarkable progress particularly over the last five decades both in the field of preclinical and clinical immunology. It would therefore be appropriate at the outset to trace the background of immunology that forms the basis for its expansion to a line at which sea and sky seem to meet.
History of Immunology Introduction to Immunology Anatomy, Physiology and Pathology of the Immune System
|
Landmarks in the History of Immunology |
1.1 HISTORY OF IMMUNOLOGY
The history of immunology goes back to 1798, when a vaccine made from cowpox was inoculated on the skin for defense against smallpox by Edward Jenner, a general practitioner of Berkeley, Glaucestershire, England (Fig. 1.1.1). The concept of this specific defense is drawn from a story, narrated by a milkmaid of her husband who was inoculated with cowpox in the arm, unwittingly, and thereafter never suffered from smallpox; and also from the experience of community herdsmen who had caught cowpox and remained unaffected by the infection, as well as, from those patients who had survived once from smallpox and never suffered again from the fatal disease. This led to the discovery of antigen, a substance that generates an antisubstance in the host with the main aim of eliminating the substance and protecting an individual from infection. This was the first record of an antigen being cross-reactive; cowpox with smallpox. It was not known to Jenner that virus causes disease.
It took almost a century more (1882) to discover that the agents, that surround us and cause infectious diseases, are pathogens. This was done by Robert Koch, a bacteriologist of Berlin, Germany. The end of the nineteenth century witnessed the demonstration of anti-substance activity in a boy aged nine years after active immunization with rabies vaccine (1888) by Louis Pasteur of Paris. The end of the nineteenth century also witnessed the demonstration of antibody activity in serum proteins called antitoxin raised against diphtheria toxin in animals. The administration of this antitoxin, in other words, the passive immunization of heterologous serum proteins with antibody activity saved the life of a little girl with terminal diphtheria at Berlin Hospital (1891) and von Behring was awarded the first Nobel Prize in Medicine (1901), in recognition for this contribution. Almost fifty years later (1950s), it was known that the cells that produce antibody and that give rise to immunity are lymphocytes leading to the explosion in active research on antibody mediated immunity and cell mediated immunity and many other functions of lymphocytes, such as destruction of virus infected cells, rejection of histo-incompatable transplanted tissues and regulation of the activity of its own and other cell types.

Fig. 1.1.1: Edward Jenner vaccinating a child(Courtesy: The Wellcom Historical Medical Museum, London)
Similarly, the history of immunogenetics, referred to as the study of the genetics of the immune system, such as Major Histocompatibility Complex (MHC), developed from transplantation reactions or rejections as a result of immune response to the transplantation antigens in allogeneic mice (Gorer, 1930) and then extended to its association with the immune response genes in immune phenomenon (Figs 1.1.2 and 1.1.3). This goes back to the concept of genetics. In 1865, Mendel practically demonstrated, in hybridization technology, that genes are particulate factors that determine traits. It would be worthwhile to recollect that the concept of inheritance in certain diseases, such as immuno-deficiencies, put forward in the past was purely based on abstract genes, i.e. recessive and dominant traits expressed respectively in homozygotes and heterozygotes. Homozygotes are individuals where the two alleles at a locus are the same; whereas heterozygotes are individuals who have two different alleles at a locus. Allele refers to the different DNA sequences, that a gene may have in a population. Almost 50 years later (1903), it was known that the factors that contain hereditary units i.e. chromosomes and the DNA, that contain millions of genes, is the genetic material. Over time, it was shown that genes lie on chromosomes, mutations of genes are physical changes, recombination of genes is caused by crossing-over and that genes code for protein.

Fig. 1.1.2: Acquired immunological tolerance: Demonstration of an induction of tolerance to make the allogeneic skin graft acceptable: tolerant strain A mouse 4 months after transplantation of CBA skin allograft). The animal had been intravenously injected within 12 hours of birth with 10 × 105 (CBA × A) (F1 A) F4 spleen cells. Hybrid cells were used to avoid runt disease, i.e. a severe from of graft-versus-host reaction(Courtesy: Leslie Brent, London)

Fig. 1.1.3: Graft-versus-host reaction (runt disease) in mice, brought about by the intravenous injection of 5 million 57 Bl spleen cells into a newborn strain A mouse. The healthy mouse is an uninjected litter mate(Courtesy: Leslie Brent, London)
Since the demonstration of differences in certain traits in the hybrids created by Mendel, it took almost a century for Crick's discovery (1953) that the structure of DNA is a double helix. The enunciation of its doctrine, knowledge of messenger RNA and triplet genetic code are the stories of post 53 era. This era also includes the sequencing of DNA, random genetic process and somatic mutation in antibody genes or even our current knowledge of the major histocompatibility complex and human genome. This also includes the impact of genomic sequencing and bioinformatic tools on our understanding towards association of genes with the immune system, hoping that histo- or haplo-identical stem cell or bone marrow transplantation, genetic engineering or genetic manipulation, gene replacement therapy or therapeutic gene cloning will also do the trick relevant to the management of diseases. There has been a revolutionary transformation from earlier technologies in immunogenetics, developed over the years to associate genetics to immune related disease into superb genomic technology.
There is a distinct shift in paradigm from immunogenetics to immunogenomics. In this process, immune related genes are found throughout the genome; four regions of candidate genes in clusters for disease susceptibility are established and these regions are MHC (Major Histocompatibility Complex), IgH (Immunoglobulin heavy chain), TcR (T cell receptor) and KIR (Killer Ig-like receptor) loci. The application of genomic techniques to genes of innate and adaptive immune response, has not only unveiled new gene 4families and shown multiple loci to contribute to the precipitation of common diseases, but it has also shown polymorphism in every aspect towards correlating genotype with immunological phenotype, the goal of any genetic analysis; as well as towards solving human diseases, the goal of any basic study in biomedical science. Whatever may be the outcome of newer molecular techniques and the use of appropriate software, our current knowledge on gene and its impact on the immune system or precipitation to certain diseases, it is yet to be appreciated that every human being is unique. Genetic disease does not stop at what we inherit from our parents. Mutation that occurs at different regions of immune related genes or elsewhere during ones lifetime may bring about changes in the traits or pattern of genetic abnormalities.
Apart from genes, certain traits or some features of human beings are determined by environmental factors, that can be seen from differences in certain characters between identical twins who are natural clones from the same egg. Thus, human beings are more than a product of their genes, signaling a great alarm to the enthusiasm on reproductive gene cloning as evident from experiments with sheep and other animals. It would be worthwhile to recollect that the mortality at infancy and that from infectious diseases is well controlled worldwide, whereas mortality from genetic abnormalities responsible for susceptibility or resistance to certain diseases is one on the rise, which may also hopefully be controlled in the near future through gene therapy, with the advent of new knowledge of human genome and development of versatile genomic techniques. However, the picture in reproductive gene cloning together with ethics attached to it, is presently alarming and there is still a long way to go in reproductive gene cloning in humans.

Fig. 1.1.4A: Eradication of smallpox as announced by the World Health Organization in 1979 following no case report of smallpox since 1975
However, as announced by the World Health Organization, the year 1979 is marked by the eradication of smallbox (Fig. 1.1.4A) and virtual elimination of many other infectious diseases, such as diphtheria, pertussis, tetanus, tuberculosis and poliomyelitis (Fig. 1.1.4B) from some parts of the world by public health measures and active immunization with potent vaccines. Immunology, thus initially developed with microbiology as a twin science to confer immunity to infection and thereafter grew, with immunogenetics or immunogenomics, to the present state by studies on man and the mouse.
1.2 INTRODUCTION TO IMMUNOLOGY
Immunology is the study of the science, as mentioned earlier that deals with the immune system. The immune system is a physiological system and a collection of cells and molecules, spread throughout the body and deployed to integrate as a whole.
The cells of the immune system are lymphocytes, phagocytes and accessory cells. These cells originate from pluripotent hemopoietic stem cells or common precursors in the bone marrow and differentiate into common lymphoid and common myeloid progenitors. Of common lymphoid, the cells of T lineage pass through the thymus and learn MHC restriction: CD4 + T Cell response to an antigen is MHC class II restricted and CD8 + T Cell is MHC class I restricted learnt at the thymus.

Fig. 1.1.4B: Virtual elimination of some other infectious disease such as diphtheria (D), whooping cough (Wc), tetanus (T), tuberculosis (Tb) and poliomyelitis (P)
The cells of B lineage pass through the stages of pro B and pre B in the bone marrow to differentiate further from immature to mature lymphocyte (Flow Chart 1.3.1). On maturation, they emigrate to the peripheral blood and tend to cluster in secondary lymphoid tissues that include spleen and lymph nodes and thereafter they recirculate between the two, i.e. peripheral blood and secondary lymphoid tissues. These cells are also found to cluster in mucosal tissues of gastrointestinal, respiratory and genitourinary tracts. The main function of these cells is to eliminate pathogens and discriminate self from nonself in order to protect the host from damage by infectious diseases and prevent the development of autoimmune disorders respectively.
The molecules of the immune system are the constitutive and inducible products of immune cells in response to stimuli. Broadly, they are antigens, antibodies, cytokines, components of complement, cluster of differentiation markers, cell surface receptor molecules and MHC proteins, in addition to transcription factors and signal transduction molecules, for expression of gene and effector function of its products, intimately connected in cross-talk in the immune system. A family of signaling receptors, referred to as Toll receptors, shown recently to play a crucial role in the induction of innate immune response in invertebrates, has also been shown to recognize invariant molecular structures of pathogens, such as lipopolysaccharide, lipoprotein, peptidoglycan and flagellin and be involved in mammalian innate immunity. Toll receptors are membrane bound receptors that do not undergo somatic mutation similar to the receptors on cell surface of the acquired or adaptive immune system, such as lymphocytes. Toll receptors are expressed on cells of the innate immune system, such as neutrophils, monocytes, macrophages, dendritic cells, NK cells or even on epithelial cells.
These cells and molecules of the immune system not only interact within the system but also interact with the cells and molecules of other physiological systems, and are tightly regulated to maintain the integrity of the body as a whole, that can be seen from the network events or the regulatory process in inflammation, antigen recognition and immune regulation.
However, the immune system, that has extensively been studied is a part of natural evolutionary process. Immunology which is primarily developed as a twin science with microbiology, and is concerned with the response of the organism to antigen derived from the pathogen and recognition of self from nonself, has infiltrated virtually every branch of medicine. It is now hard to find a single disease, which does not have an element of immunology. It is also hard to find a specialty in biomedical sciences, which does not have a link with immunology. These are immunophysiology, immunochemistry, immunopharmacology, microbial immunology, immunogenetics, immunoneuroendocrinology, immunohematology, immunopathology and immunobiology. The branches of immunobiology encompass transplantation immunology, histocompatibility testing and comparative immunology. Numerous attempts are being made to translate the outstanding academic excellence achieved over the last 50 years in basic science into practice, in other words to link basic immunology to clinical immunology for the diagnosis and treatment of infection and immunological diseases, such as immunodeficiencies, hypersensitivities, auto-immunities and malignancies (Flow Chart 1.3.2).
Of great relevance of immunology to humanity is the success in eradication of smallpox by vaccines. There has been also a great success in the prevention and elimination of many other infectious diseases. Safe blood transfusion and better understanding of transplantation immunity are other success stories in immunology. With better understanding of the MHC and its relationship with the immune reactions, together with newer immunosuppression regimes that include cyclosporin A and FK 506 that act synergistically to block T cell or IL-2 responses, the peripheral aspect of transplantation biology has grown enormously. Transplantation of organs or tissues, such as kidney in patients with end-stage renal disease, bone marrow transplants in patients with certain genetic disorders or malignant diseases, has become a common surgical practice. There have been also transplants of heart, heart-lung, lung, liver, pancreas, bone, cartilage, brain, anterior chamber of the eye and cornea. In this, cornea is avascular where as bone and artery are largely acellular. Over and above, cartilage, tendon, major blood vessels and even fetus are privileged tissues, due to lack of expression or low content of MHC molecules; whereas womb, brain and anterior chamber of the eye are privileged sites that tolerate grafting without sensitization of the recipient.
It is thus imperative for everyone in this field, even the non-expert, to comprehend the basic concepts and 6overall picture of immunology, which has undergone an evolution of stellar proportions over the last two hundred years. There is also enough evidence to suggest that the resistance of an individual to disease improves with serene state of mind and mental peace.
It would take too long to cover the subject in depth here and hence, various models, such as allo-recognition and state of mind with disease are dealt with. There are many excellent texts and reviews that will help readers not only to overcome difficulties in understanding but also to take students of medical sciences further. This overview is just to provide key elements and focus in brief the strategy of the immune system. Those who are already familiar with the topic may find it easy to understand the fascinating concepts of immunology and expand further from the building blocks drawn here to reach, step by step, the other end of the contemporary science. It may appear to be an attempt to duplicate the efforts of others, but is not. It aims to make the complexities of the learning process simpler and improve on the current level of understanding.
In this overview, I have first defined immunity and then classified it, with an introductory account of the elements of innate and acquired immunity. This overview also narrates, in some detail, the strategy of the immune system, the components of which include cells and molecules, development of reactive lymphocytes, elimination of anti-self cells through programmed cell death or apoptosis and generation of anti-nonself cells in the antigen independent process by genetic recombination. The components of the immune system also include cells that recognize antigens and integrate signals that they perceive in their environment in the antigen dependent process by genetic reprogramming. I have followed that by a brief outline of immune response leading to immunity to infection and disease of immunity. I have also followed that by a quick reference to the immunology of certain diseases, a brief note on immunological tests, a bird's eye view of molecular techniques, an outline on immunological investigations relevant to the management of the patient and finally an immunology at a glance. The anatomy, physiology and pathology of the immune system includes a major site of immune reactivity, the basic unit of all living matter and the immune response to an antigen along with a note on landmarks in immunology and turning points in history.
1.3 ANATOMY, PHYSIOLOGY AND PATHOLOGY OF THE IMMUNE SYSTEM
- A major site of immune reactivity
- The basic unit of living matter and network interaction
- The immune response to an antigen and the genetics of the immune system
- The anatomy of the immune response
- The physiology and pathology of the immune response.
A MAJOR SITE OF IMMUNE REACTIVITY
A major site of immune reactivity is referred to as lymphoid tissues, including aggregates of lymphocytes of mucosal immune system.
Lymphoid tissues of the immune system, as mentioned earlier, are traditionally grouped under primary and secondary (Fig. 1.3.1). The bone marrow and thymus constitute the primary or central lymphoid tissues and the solitary spleen and multiple lymph nodes, widely distributed all over the body constitute the secondary or peripheral lymphoid tissues. The thymus, developmentally emerged from the third and fourth pharyngeal pouches, is bilobed, located retrosternally and composed of epithelial cells, lymphoblasts or lymphocytes with MHC restriction. Whereas the bone marrow is composed of vascular adipose tissue and precursor cells of lympho-myelo-harmopoietic series, the spleen and lymph node act broadly as a filter, an important site for phagocytosis and the initiation of immune responses to the antigens.
Immune response is defined as a phenomenon that occurs in a suitable host, man or animal, when foreign antigen enters the body in soluble or particulate form and interacts with cells of the myeloid and lymphoid series. Cells of the myeloid series, such as macrophages, process the antigenic materials and present the determinants directly or through antigen presenting cells to the cells of the lymphoid series which are immunologically competent, such as lymphocytes.7
In response to an antigen, the naive lymphocytes pass through the stage of effector and memory cells, such as antibody secreting effector plasma cells, lymphokine producing CD4+ helper and CD8+ cytotoxic or cytolytic effector T cells, memory B cells and memory T cells. Accessory cells of the myeloid series, such as dendritic cells are specialized cells which may be located in the common portals of entry of microbes, such as skin and respiratory, gastrointestinal and genitourinary tracts. These specialized cells may capture antigenic substances derived from the microbes and transport them to the peripheral lymphoid tissues where antigen recognition takes place and lymphocytes are transformed into antibody secreting effector plasma cells and lymphokine producing CD4+ and CD8+ effector T cells, in addition to the generation of a section of primed lymphocytes into memory B and T cells. Though the dendritic cells are referred to as professional antigen presenting cells to the naive T cells for activation and the release of IL-4 for differentiation of antigenically activated B cells into effector plasma cells, the cell types for capture of non-self antigen and presentation to the B cells are not known. In contrast to the T cells, the B cells may directly recognize microbial antigens, such as polysaccharides to produce antibodies and eliminate the microbes. However, it was thought that a type of dendritic cells called follicular dendritic cells, that reside in the germinal center of the lymphoid tissues and that do not present antigen to the T cells, may display antigen to the B cells for differentiation of activated B cells into plasma cells.
It was first shown in birds, in particular in chickens, that an early removal of the thymus affects cellular response; whereas an early removal of bursa of fabricius affects humoral reponse. Though there is no true organised tissue in mammals to represent the bursa, the yolk sac and foetal liver and the bone marrow itself and particularly peripheral lymphoid tissues themselves in adults are presently considered as possible sites to the maturation of B-lymphocytes.
|
|
Yolk Sac and Foetal Liver
In mammals, the yolk sac is the earliest and the foetal liver is the next site of the precursor lymphocytes; in which major haempoiesis and lymphopoiesis take place that produce all red and white cells of the blood.
Bone Marrow
Bone marrow is composed of (a) vascular-adipose tissue and (b) lympho-haemopoietic tissue. Each of these two groups of tissues approximately weight about one and a half kilogram in an average man. While the vascular-adipose tissues provide nutrients and removes wastes of metabolically active cells, the lympho-haemopoietic tissues provide with all cells of the blood and the immune system. Though the exact sites for lymphopoiesis are yet to be clearly demarcated from the sites for haemopoiesis in the bone marrow, the lympho-haemopoietic system is further divided into four main series of tissues: (a) erythroid, and (b) megakaryocyte (Fig. 1.3.2), (c) myeloid, (d) lymphoid, that respectively give rise to four distinct group of cells: erythrocytes, platelets, lymphocytes and monocytes or granulocytes. While the lymphocytes and monocytes or granulocytes are directly involved in the immune response, the erythrocytes and platelets are indirectly involved as target cells of the immune response.
However stem cells derived from lymphoid series in the bone marrow (Fig. 1.3.3) and destined to the pool of B-lymphocytes pass into the central vein for their discharge into the circulation via the blood to the bursa of fabricius in birds or equivalent in mammals; whereas stem cells derived from lymphoid series and destined to the pool of T-lymphocytes pass via the blood to the thymus, to acquire competence or restore normal function in the host in the absence of bursa equivalent site and thymus.8

Fig. 1.3.1: Anatomical distribution of central and peripheral lymphoid tissues. Primary and secondary lymphoid tissues(Courtesy: JB and DM)
Primary | — | bone marrow and thymus |
Secondary | — | lymph nodes: cervical, axillary, mucosal and inguinal |
— | spleen | |
• A site of antigenic stimulation of lymphocyte during its passage through lymphoid tissues leading to | ||
↓ | ||
• Inflammatory or immune response and deployment of its products that include cells and molecules of innate and acquired immunity constituting cellular and molecular barriers of internal tissues. | ||
• Lymphoid tissues are spread all over the body and connected through lymphatic and circulatory network. | ||
Thymus
The thymus is developmentally emerged from the third and fourth pharyngeal pouches. It is usually a bilobed (Fig. 1.3.4) lymphoid organ which may be multilobed in some species. It is central to immunological functions. It weighs about 15 gm in mice at birth and reaches its largest size upto 35 gm at puberty and thereafter gradually wanes until it becomes fully atrophied.
The thymus is enclosed by capsule and divided by trabeculae into a number of lobule has outer cortical and inner medullary areas. These areas contain two major groups of cells. They are called as fixed tissue cells and migratory lymphoid cells. The fixed tissue cells, are epithelial and recticular cells. They are respectively derived from mesoderm-endoderm of the third pharyngeal pouch and spread throughout the thymus. The migratory lymphoid cells are derived from the foetal liver and bone marrow at different stages of development of the thymus via the blood, which aggregate within the framework of epithelial-reticular cells and are pure population of lymphocytes. In the cortical area, the epithelial-reticular cells stain dark, and are densely populated with actively dividing lymphocytes of various shapes and sizes. These rapidly dividing cells are immunologically immature and physically sensitive to cortisone. Ninety percent of these self reactive cells are small, 9 percent medium and 1 percent large, but the reason for high turnover rate and the formation of Hassall's capsules as aggregates of various cell types is still obscure.

Fig. 1.3.2: Bone marrow multipotent stem cells differentiated into cells of lymphoid, erythroid, megakaryocyte and myeloid series
Pyknotic cell with multiple nuclei as a sign of cell death is also a common feature in the cortical area.

A number of cell divisions, within the thymus, perhaps 6-8, have been shown to exist before the precursor cells become immigrants in the periphery. The epithelial-reticular cells with dendritic processes, which express gene products or surface Ia+antigen in the medullary areas, are on the other hand, thinly populated with immunologically mature and physiologically cortisone-resistant lymphocytes, which may have undergone some differentiation in the bone marrow under the influence of thymic hormone. Otherwise we know very little about the nature of the epithelial environment of the thymus and its relationship to cellular differentiations. We are also quite ignorant of the molecular mechanism involved in cellular differentiation, though a number of soluble factors extracted from thymus have been shown to stimulate the maturation of T-lymphocytes. The epithelial cells of the thymus are, however, thought to produce at least one, if not more, hormone known as thymosin which is considered to be important in the maturation of the T-lymphocytes. Though the thymus in the mouse contains almost exclusively cells of T series, there may be a small number of B-series in some animals that derive from bone marrow and are distinguishable by the presence of thy1 antigen on T-lymphocytes and its absence on B-lymphocytes.
Furthermore, it is yet to be made clear, whether all T-lymphocytes pass to the medulla from the cortex before leaving for peripheral circulation or some population leave directly from the cortex without passing through the medulla which may be the site of maturation or immuno-competence. The relationship, between the different stages of the differentiation of lymphocytes within the thymus and the development of cancer or ageing, is still a matter of speculation.

Fig. 1.3.4: Thymus showing characteristic features of each lobule with respect to the distribution of epithelial cell, reticular cell and thymocyte at different stages of their development
The evidence of thymic abnormalities and immune deficiency states demonstrated by Good and his associates in man, particularly in children, has led to the suggestion that the thymus is intimately associated with the function of the immune system. This has experimentally been shown by Miller and his associates in neonatally thymectomized mice, in contrast to mice thymectomized in adult combined with or without irradiation.
Effect of Neonatal Thymectomy
Neonatal thymectomy in mice has a profound effects on the immune system that are characterized by:
- Deficiency in their capacity to respond to various antigens, such as Salmonella typhi and killed influenza virus; heterologous red blood cells and transplantation antigen;
- Incapability of initiating graft-versus-host reaction;
- Inability to respond to T-dependent antigens and thereby to give rise to cell-mediated immune response;
- Tendency to develop a wasting syndrome characterized by diarrhoea, loss of weight, and susceptibility to infection such as salmonella, brucella, acid fast bacilli, listeria, fungi, vaccinia, measles and chickenpox that simulate to a situation in which a new born develop the above features following allogeneic or foreign spleen cell transfer; and
- Tendency to develop lymphoid aplasia, low peripheral blood T and B lymphocyte level, a low output of thoracic duct lymphocytes and specific depletion of T lymphocytes in T-dependent areas of spleen and lymph nodes.
The intimate association of thymus with the immune system has later been shown in certain strain of hairless mice which possess thymus sacs lacking lymphocytes. This strain of mice is known as nude mice. It has further been shown that the defect in nude mice lies not in the precursor stem cells but in the process of maturation that takes place in the thymus. This has been shown by the transplantation of bone marrow stem cells from nude mice into normal mice and the demonstration of repopulation in the thymus that was irradiated prior to stem cell transplantation. It means, nude mice are incapable of mounting T-cell or cell-mediated immune responses.
Effects of Adult Thymectomy
Adults thymectomy has little effect as compared to thymectomy at neonatal life in mice. However, some effects can be demonstrated if thymectomy is combined with lethal irradiation with 850r which totally destroys the ability of lymphocytes to divide and thereby their ability to respond to antigens. The administration of syngeneic bone marrow cells (as a therapeutic measure to keep them alive) can however, restore the ability of antibody production by lymphocytes in thymectomised and lethally x-irradiated adult animals. A normal condition can be achieved in the situation by an additional implantation of a thymus graft which perhaps function similar to a mechanism seen in endocrine organ transplantation.
Syngeneic bone marrow alone is not immunologically fully effective in adult animals who have had both thymectomy and lethal irradiation. This suggests that thymus exerts a significant influence on bone marrow stem cells for the immune response that is, thymus is required for its recovery. It has further been shown that a state of tolerance induced in an adult may be prolonged when thymectomy is also done, indicating that continuous supply of antigen sensitive cells, perhaps T-cells, are essential to terminate tolerance. Although adult mice following thymectomy and x-irradiation with 350r can produce small amounts of antibody and take skin homograft for a longer period than usual, such animals recover from suppression by about four to six weeks.
Thymic Hormone
Though the thymus still remains to be shown as an endocrine organ, yet the ‘soluble factors’ isolated from the primary lymphoid epithelial tissues or thymus of a variety of species are known as thymic hormones. A number of such hormones and their ‘synthetic analogues’ which mimic the role of natural products of the thymus are peptide in nature with 1000 to 8000 mol. wt. Some of these are known as thymosin (2000-5000 mol. wt.), thymopoietin I and II, (6000 mol. wt.), ubiquitin, (9000 mol. wt.), thymic humoral factor in short THF (3000 mol. wt.) and facteur thymic serique, in short FTS. Of these, FTS was initially isolated from pig serum and recently from thymus. FTS disappears from the serum with age and at the onset of autoimmune disorders and after thymectomy. FTS interacts with T-cell receptors and enhances the formation of cAMP. It has also been shown that FTS stimulate suppressor populations which are dependent on high concentration of the hormone. Studies on hormones of thymic origin were of great interest in order to understand the regulation of T-cell as well as to find a solution of the management of immunodeficiencies. In spite of some insight, none of the factors have been shown to substitute the role of intact thymus and correct the T-cell deficiency states in certain clinical situations.
Action of Thymus
Theoretically there are mainly two possible ways, cellular or humoral, by which the intact thymus or thymus graft can play a major role in facilitating the development of the property of immunological responsiveness. That is, the thymus plays its role either by commitment to a population of cells, which become a part of the immunological apparatus. There is an evidence to suggest that the deficiency of T-cells perdisposes an inidividual to the infection with intracellular organism and the development of cancer. Or thymus plays its role by secreting a humoral or hormonal factor similar to a mechanism involved in an endocrine organ.
Bursa of Fabricius
In birds, there are three central lymphoid tissues as compared to two in animal and man. The bursa of fabricius is the third in birds over and above marrow and thymus common to bird and animal. Similar to the thymus, Bursa reaches its largest size in early life. It is derived from the gut epithelium but restricted to the species of birds (Fig. 1.3.5). Though various lymphoid tissues, such as appendix and tonsils, have been claimed, there is perhaps no analogue of bursa of fabricius in mammals.11

Otherwise, similar to the thymus, the bursa consists of fixed and migratory cells in cortical as well as in medullary areas. The cortex is the site of proliferation and the medulla is the site of colonisation of immuno-competent B-lymphocytes for onward transportation or immigration to the peripheral lymph node and circulation.
Effects of Neonatal Bursectomy
Neonatal bursectomy in chickens, with irradiation, which eliminate the capacity of the existing lymphocytes to divide, has shown that there is a gross reduction in humoral antibody production; whereas cell-mediated immunity is intact. In contrast, cellular immunity is grossly impaired by neonatal thymectomy in mice, but there is always some effect on antibody production because the production of antibodies to some antigens depend on T-lymphocytes.
Peyer's Patches
Peyer's patches are aggregates of a population of lymphoid cells in between the inner mucus membrane and the outer muscular layer of the intestinal tract. They do not have afferent lymphatics, but only efferent routes. One important feature of these patches is the presence of large lymphoid follicles with highly active germinal centres, perhaps because of constant stimulus from breakdown products of micro-organisms in the gut.
Otherwise they are very similar to lymph nodes in structure. The intestinal wall contains lymphocytes which are capable of producing IgA as a first specific defence against invasion by local micro-organisms in and around the gut.
Although the bursa equivalent organ of lymphoepithelial structures in mice and man are not well defined, the removal or Peyer's patches from the intestine of rabbits and an atropic disease of the human intestine, have shown that gut-associated lymphoid tissues in man may be equivalent to the bursa of chicken.
Spleen
The spleen (Fig. 1.3.6) is one of the most important sources of antibody producing cells. From the evolutionary point of view, it is older than lymph nodes. The spleen is enclosed by a capsule and divided into many compartments by trabecules. The lymphoid tissues in the spleen can be divided into two distinct areas held by supporting tissues and these areas are known as white pulp and red pulp.
The white pulp is situated around the branches of the splenic artery. The lymphatic nodules that surround the artery are termed as malpighian bodies. Malpighian follicles in the white pulp are analogous to follicles of the lymph nodes and Peyer's patches.

Fig. 1.3.6: Spleen showing red pulp, white pulp, supporting tissues together with germinal central arteriole and thymus dependent area and periarteriolar space
That is, these areas are similar to the cortex of the lymph node. The regions immediately adjacent to the arteriole in the white pulp of the spleen are considered as thymus-dependent areas similar to paracortical region of lymph node. The red pulp on the other hand is composed of sinuses and medullary cords which constitute about 50 percent of the spleen. It plays an important role in engulfing damaged red cells and destroying erythrocytes. Although the exact pathway of circulating lymphocytes in the spleen is not known, the lymphocytes are thought to enter the arterioles and interact with antigen. These may perhaps leave the white pulp through capillaries and enter into the red pulp. These activated cells after passing through the sinuses reach the venules and thereby the veins in one direction, and the surrounding tissues within the spleen in another. However, the spleen is associated with multifarious activities.
Lymph Node
The lymph nodes (Fig. 1.3.7) that are widely distributed throughout the body, are the main source of antibodies. The lymph node is roughly spherical in shape and is structurally less complex than the spleen. The structure of the lymph node in man and animal is almost similar. Like the spleen, the lymph node is encapsulated and divided by trabeculae, which extend from the cortex to the medulla. The space covered by the capsule is filled up with lymph termed as circular sinus. The lymph has two main pathways; one being directed into the medulla through the trabecular sinuses, and the other directly through the outer lining of the circular sinus into the cortex. Unlike the spleen, the antigen enters into the circular sinus of the lymph node through afferent lymphatics. The sinus communicates with the afferent and efferent lymphatics through which the lymph flows from different parts of the body and then passes into the main lymphatic trunks and veins.
The cortex of the lymph node is filled up with lymphocytes. The densely packed collection of these cells constitutes an inconspicuous nodule termed as primary follicle all over the surface of the cortex. In this, there are special veins termed as post capillary venules, which are lined with thick cubical endothelial cells. The special feature of these venules is that the recirculation pools of lymphocytes pass through the cytoplasm of these lining cells rather than through intercellular space or in between two cells through which other cell types pass.

Fig. 1.3.7: Lymph node section showing cortical, paracortical and medullary areas together with sinuses and supporting tissues
However, the lymphocytes grouped together in the form of follicles in the cortex are surrounded by reticular cells and macrophages with their cytoplasmic projections (Fig. 1.3.7). The medulla is made up of cords that extend from the cortex and open into the sinuses which are lined by enormous number of macrophages which filter the infective or foreign agents and processing of the antigenic substances for presentation to the lymphocytes. Macrophages are also found to be scattered throughout the gland, many being in medullary areas, together with plasma cells and their precursors in the cortico-medullary junction. Though the T-cells in general tend to reside in medullary and B-cells in the cortical regions of the lymph nodes, the paracortical areas are predominantly packed with T-cells and both cortical as well as medullary areas are packed with B-cells. The areas that contain predominantly T-cells are known as paracortical or thymus dependent areas; whereas the areas that contain predominantly B-cells are known as cortical or thymus independent areas. These areas undergo certain changes after infection on recognition of antigen; the virgin lymphocytes may be transformed in about 18 to 24 hours into blast like cells. The number of these cells gradually increases in 2 to 3 days and this along with recruitment of new cells in the vicinity constitute 8 to 16 small inconspicuous areas in the lymphoid tissue known as primary follicles. The building up of accumulation of such cells in large number in an area that constitute the secondary follicles is termed as germinal center (Fig. 1.3.8).13

The cells, of these centers engaged actively in antibody formation, are localized predominantly in corticomedullary regions of the lymph nodes and red pulp of the spleen. Though the ultimate fate of all the cells born in the germinal center has not been possible to trace, about 100 to 1000 transformed cells, well equipped with important constituents, necessary for cell division are enough to constitute the center. There is no fixed number of mitotic divisions for a newly transformed cell. It can vary from 6 to 8 sequential divisions at 8 to 10 hours interval under antigenic stimulation (Fig. 1.3.9). In other words, these cells divide twice or thrice a day. Some of these die in 2 to 3 days and others are transformed into effective cells and memory cells. While the former live for a few days to weeks after being transformed into plasma cell, the latter live for months to years.
The antibody forming plasma cells (Fig. 1.3.10) measure between 10 to 20 µ. Mature plasma cell has an eccentric nucleus with densely packed chromatin, often arranged as a clock face. In contrast, the primitive cell has a larger centrally situated nucleus with finely dispersed chromatin. The cytoplasm of a plasma cell is well equipped with ribosomes bound to the endoplasmic reticulum. This is associated with protein synthesis, and capable of secreting antibodies, the aggregates of which in lymphoid tissues are known as Russel bodies.
The division of cells on antigenic or mitogenic stimulation as stated above can be detected by incorporating radioactive precursors thymidine, since cells can never be expected to divide without proper duplication of DNA molecule, indicating that the number of granules incorporated in a particular cell may give an idea about the number of division undergone by the cell. It means, the number of cell divisions may approximately be determined by counting the number of granules in each daughter cell.

Fig. 1.3.9: Plasma cell showing various structures: nucleolus, nucleus, golgi apparatus, endoplasmic reticulum with ribosomes, mitochondria and Russell body
However, the synthesis of DNA, RNA and protein differ in different stages of the development of these cells and the rate of synthesis of these molecules is usually high at an early stage. As these cells get fully matured, the RNA synthesis slows down, the DNA synthesis stops and the protein synthesis is predominantly directed to antibody formation.

Fig. 1.3.10: Antibody formation: synthesis of light and heavy polypeptide chains that lead to the formation of antibody molecule

Fig. 1.3.11: Bone marrow precursor cells pass through thymus and bursa in birds and recirculate between peripheral blood and lymphoid tissues
The m-RNA molecules are very stable in long-lived antibody-forming cells. The coded instruction from DNA is carried by m-RNA to ribosomes which are capable of reading the RNA message for the synthesis of light and heavy chain are shown by radioisotope technique. The light and heavy chains thus formed are assembled to make an immunoglobulin molecule of one specificity (Fig. 1.3.11). This molecule, undergoes glycosylation, that requires carbohydrate moiety, which is derived from the cell membrane before it is secreted into the extracellular fluid.
Actively antibody-secreting plasma cells do not undergo further division. These cells are exhausted in 2 to 4 days and finally eliminated from the system. However, the antibodies thus formed and secreted in extracellular fluid may combine with stimulating antigen and form immune complexes predominantly to eliminate the foreign substances or particularly infective agents and partly to regulate the immune response. The immune-complexes thus formed can also cause damage to the tissues under certain circumstances. Whatever may be the consequences of the immune response, anatomically stem cells derived from the bone marrow enter as pre T-cells and pre B-cells respectively into thymus and bursa where maturation of the cells take place for further transmission to the peripheral circulation, spleen and lymph nodes (Fig. 1.3.12) for activation in response to the antigen.
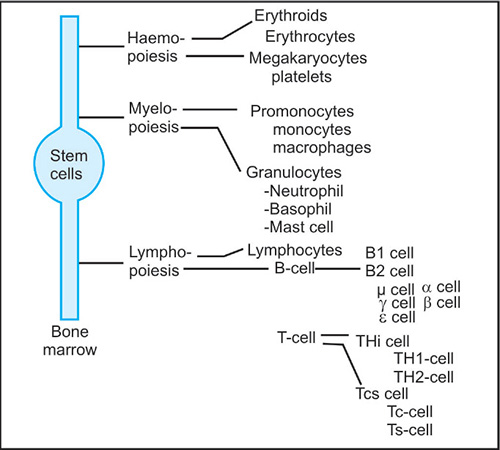
Fig. 1.3.12: Cells of lympho-myeloid series that participate in various innate and acquired function
Mucous Associated Lymphoid Tissues (MALT)
MALT are aggregates of lymphocytes near mucosal surface of respiratory, gastrointestinal and genitourinary tracts. There is local synthesis of secretory IgA antibodies which bind to microorganisms, bacteria or viruses, and prevent their adherence to the cells of the mucous membrane, and thereby their multiplication and entry into the body.
Secretory IgA antibodies are also present in tears and salivary secretions. There is also local T Cell activity and synthesis of IgE antibody, in which IL-4 release by T Cell helps in differentiation of activated B cell switch over to IgE producing plasma cells in a linked recognition. If there is any breach in the surface barriers, IgE antibodies bound or coated to mast cells come to the rescue and provide the next line of defence by degranulation, release of histamine and passage of cells, antibody and complement proteins from the circulation to the site of infection.15
Cells that Derive from Lympho-myelo-Haemopoietic Stem Cells of the Bone Marrow
Cellular aspects of the immune response include cells that derive from lympho-myelo-haemopoietic stem cells of the bone marrow. These stem cells are multipotent that has experimentally been shown by (a) cell transfer in vivo, (b) cell cloning in vitro, and (c) chromosomal analysis in irradiated mice. These stem cells, that form the basic unit of all living matter, are undifferentiated precursor cells, that can in due course broadly be differentiated into cells (Fig. 1.3.12) of (a) haemopoietic series, (b) myeloid series and (c) lymphoid series under the influence of certain hormones.
THE BASIC UNIT OF LIVING MATTER AND NETWORK INTERACTION
The basic unit of all living matter is the cell that constitute the body; whereas the molecule is the basic unit of network interaction in which the molecule may either be free in the extracellular body fluid or bound on to lipid bilayer of the cell membrane as cell surface markers such as receptor or cluster of differentiation or CD molecules that constitute the anatomy, physiology and pathology of the immune system. These features are common for all physiological systems.

Fig. 1.3.13: The basic unit of all living matter is cell that passes through four phases: mitotic (M) phase, gap 1 (G1) phase, synthetic (S) phase and gap 2 (G2) phase. The cell, after mitosis or division (1-2 hr), enters into a quiescent phase G1 (4-5 hr). It is followed by uncoiling of the chromosome and synthesis of DNA (7-8 hr) leading to the second phase of quiescent or cessation of DNA synthesis or G2 (2-3 hr) and ending with coiling of chromosome
- Each of these cells passes through a life cycle and runs on internal or external activation in four phases (Fig. 1.3.13).
- Each of these cells comprises of a nucleus, cytoplasm and membrane (Fig. 1.3.14).
- Cytoplasm of the cell is constituted by several organelles, such as ribosome, endoplasmic reticulum and Golgi apparatus. They are involved in protein synthesis as per codon or genetic code and channelization into the cytoplasm for further transport into the lipid bilayer of the cell membrane to form cell surface receptor and constitute marker or release in extracellular fluid.
- Cytoplasm of the cell also contains other organelles such as vacuoles as a store house of nutrients, lysosomes as reservoirs of enzymes to metabolize the nutrients and mitochondria as a powerhouse of energy.
- Nucleus of the cell similarly contains nucleolus, and all the genes of the whole organism in the form of a double-helix or strand called DNA and each of these genes contains the coded information for one protein, that means, one gene and one protein except those like immunoglobulin that requires more than one gene for the synthesis of one antibody protein.
- DNA is a double-helix or double stranded molecule, as mentioned earlier, that consists of a sugar (deoxyribose), a phosphate group and four nitrogenous bases: adenine (A), cytosine (C), guanine (G) and thymine (T).A exclusively pairs with T and C pairs with G to constitute A-T and C-G base pair. DNA bases encode messenger RNA (mRNA) which in turn encodes amino acid sequences. Whereas RNA is a single stranded molecule that consists of a sugar (ribose), a phosphate group and four nitrogenous bases: adenine (A), cytosine (C), guanine (G) and uracil (U). There are three basic forms of RNA and they are messenger RNA (mRNA), ribosomal RNA (rRNA) and transfer RNA (tRNA). Nucleotide is a basic unit of DNA or RNA, consisting of one sugar (deoxyribose for DNA and ribose in the case of RNA), one phosphate group and one nitrogenous base.

On activation from inside or outside the cell, a copy of a particular gene contained within the double strand DNA molecule is made into RNA in the nucleus and the process of copying DNA information into RNA for onward transmission is termed as transcription. Genetic code is the combinations of mRNA codons that specify individual amino acid (Table 1.3.1) and codon is a group of three mRNA bases, each of which specifies an amino acid when translated. It means codon is a triplet code, a sequence of 3 bases. There are 64 codons to code for 20 amino acids that form conventional proteins and super family antigens. However, the coding information that remain in the portion of nucleotide sequences of gene, known as exons, can often be interrupted by stretches of the adjacent portions of nucleotide sequence, known as introns that lack coding function but share 90 percent of DNA indicating what an enormous amount of excess DNA we all possess (Fig. 1.3.15).
Since the primary transcript RNA in the nucleus is a true copy of both exons and introns, the latter is snipped out of the RNA to reflect only true copy of the exons as a functional unit. It is at this stage that the RNA is ready to make move from nucleus to the cytoplasm as messenger RNA, in short mRNA, that attaches to ribosomes and codes for synthesis of different amino acids to form protein; and the process of coding by mRNA to synthesize different amino acids and form a particular protein is termed as translation. It means, the formation of protein involves DNA mRNA transcription and mRNA protein translation. The proteins thus formed pass through the channel of the endoplasmic reticulum into the cytoplasm and thereafter are taken up by the golgi apparatus to the lipid bilayer of the membrane that may form receptors on the cell surface. The receptors (surface immunoglobulin) (SIg) in the case of B cells may be produced on activation from inside the immuno-competent cells or be secretory in the extracellular fluid on activation from outside the immuno-competent cell by the antigen. Similarly such proteins or glycoproteins may also be produced by other cells: such as interleukin-1 by monocytes and MHC class II antigen by antigen presenting cells that participate in a network of complex interaction in antibody and cell mediated immune response.
There are more than one kind of cells in man. Each of the cells produces different molecules and thereby performs different functions, for example nerve cells of the brain produce neuropeptides to act as neurotransmitter, β-cells of the pancreas produce insulin to monitor blood sugar and B lymphocyte of the immune system produce antibodies to play role in defense and regulation of the immune response.

Fig. 1.3.15: Cell, the basic unit of all living matter, contains nucleus where DNA exists as double-helix. On activation of gene, the two strands of DNA separate and a step is followed in which mRNA (a true copy of DNA) is made. The mRNA, that contains stretches of both functional exons and non functional introns, is processed inside the nucleus and after intron sequences are cut out to move into the cytoplasm. The ribosomes attach to the mRNA and each triplet of their bases coding for one particular amino acid progressively assembled to form a protein chain. When the ribosome comes to the end of the mRNA, it falls off and the protein is released
It also shows that the molecules that participate in an array of secondary events involve a number of other effector cells and their byproducts which soon become embodied in a network of complex immune response and non-immune reactions, such as inflammation indicating their principal role in more general term rather than in a specific or precise biological sense. However, the phenomena, depending on the cell types and their products, together with genetic background in a particular situation, influence the dynamics of the innate and acquired immune reaction. Implications include in clinical medicine resistance to infection, pathological consequences or diseases of immunity and immunological diagnosis relevant to the management of the patients.
THE IMMUNE RESPONSE TO AN ANTIGEN AND THE GENETICS OF THE IMMUNE SYSTEM
Immune response in an acquired immune reaction is otherwise a phenomenon of lymphocyte differentiation into effector and memory cells in response to an antigen (Fig. 1.3.16). Antigen receptor on B-lymphocyte is different from antigen receptor on T-lymphocyte. B-lymphocytes usually recognize and bind free antigen through membrane immunoglobulin receptor for differentiation of cells and production of antibodies in T-independent immune recognition; whereas T-lymphocytes usually recognize processed antigen in association with MHC molecules through TCR-CD3 receptor for activation of cells and release of lymphokines in order to help antigenically activated B-lymphocytes for differentiation into plasma cells and production of antibodies in T-dependent linked recognition through antigen presenting cells e.g., macrophage.18

Lymphocytes have enough cell surface antigen receptors to recognize any biological molecules. The diversity of these antigen receptors is generated during lymphocyte development and before cells encounter an antigenic molecule. It means, the immune system does not know in advance what it will be asked to recognize.
In such situation, the immune system must have a huge number of structurally diverse lymphocytes, so that a few cells can bind and respond to any antigen and thereby the humoral immune system can generate billions of structurally distinct antibodies, the mechanism of which is complex, particularly in view of more than one gene for one antibody rule and the recent estimates of 35,000–50,000 genes in human genome. This refers to all of the DNA in an organism. However, a number of mechanisms for generating antibody diversity have been considered to be operative and they are germline immunoglobulin genes and somatic VDJ (variable, diversity and joining) recombination together with mutation and hypermutation of genes. These may be superadded by a number of others, such as variations of a few nucleotide sequences (deletion or insertion) and thereby a few amino acids at joining sites as well as random combination of heavy and light chain during assembly of immunoglobulin.
Immunogenetics, as mentioned earlier, is the study of the genetic basis of the immune system in health and disease. Mendel (1865) laid down the foundation of genetics through his discoveries of inheritance. Rediscoveries of his principles occurred through ABO blood group by Landsteiner (1900s) and histocompatibility by Gorer (1930s). The term gene is coined to denote the basic unit of heredity by Johannsen (1909) and these genes are composed of deoxyribonucleic acid or DNA (Avery 1944). In 1953, Watson and Crick described the physical structure of DNA as a double helix. A double stranded DNA consists of sugar phosphate backbone and four nitrogenous bases: adenine (A), cytosine (C), guanine (G) and thymine (T). A and G are purine bases in both DNA and RNA; whereas C and T are pyrimidine bases in DNA and C and uracil (U) in RNA. As a rule, A pairs with T (A-T) and C pairs with G (C-G) that constitute A-T or C-G as base pair in DNA. There are more than 3.5 billion base pairs. This formed the basis of molecular genetics and helped in the study of the structure and function of the genes at the molecular level. Nucleotide is a basic unit of DNA or RNA, consisting of one deoxyribose (or ribose in the case of RNA), one phosphate group and one nitrogenous base. DNA bases or portions of genes that encode messenger RNA (mRNA) and in turn encode amino acids sequences are called exons and the DNA sequence found between two exons is intron.
The physical location of DNA is the chromosome (1956), a thread-like chromatin body in the nucleus of cell. The mitochondria are cytoplasmic organelles that have their own unique DNA. In human, there are 46 chromosomes, i.e. 23 pairs of chromosomes: 22 pairs are autosomes and one pair is the sex chromosome (XX or XY). X-linked refers to genes that are located on the X-chromosome, whereas a transmission exclusively from father to son refers Y-linked. Since 1960, there has been a significant technological development and over the last two decades, a large number of genes have been mapped. In Human Genome Project (1990–2003), each human is estimated to have 35,000–50,000 different genes in the genome that refers to all of the DNA in an organism. Any alterations in the base pair or gene sequence or in chromosome, brings about genetic disorders in which entire chromosome may be missed or duplicated respectively causing Down syndrome and Turner syndrome. Single genes may be altered causing single gene disease (mendelian) such as cystic fibrosis, sickle cell anaemia and haemophilia. Disorders may result from combination of multiple genes and environment such as life style or infection causing diabetes and heart disease. However, disorders of the immune system, such as primary immunodeficiencies are usually directly caused by defects in the genes. They result in insufficiencies in the molecular components or cellular elements of the immune system and manifest clinically as susceptibility to infection.19
THE ANATOMY OF THE IMMUNE RESPONSE
The immune system, as mentioned earlier, is a collection of cells and molecules (Flow chart 1.1). The cells of the immune system that constitute the anatomy of the immune response consist of: (a) phagocytic cells of innate immunity, (b) some accessory cells having specialized glycoproteins as integral cell surface molecules or membrane proteins and (c) lymphocytic cells of acquired immunity, that constitute the lymphoid tissues, traditionally divided into primary or central and secondary or peripheral lymphoid tissues (see strategy of the immune system). The examples of primary lymphoid tissues are bone marrow and thymus; whereas examples of secondary lymphoid tissues are spleen and lymph nodes (LN), which are of two types: encapsulated and unencapsulated. The encapsulated lymph nodes, spread throughout the body, are structurally or architecturally well equipped to give rise to the production of virtually all classes of antibodies; whereas the unencapsulated lymphoid tissues, localized as aggregate or in diffuse form and restricted to subepithelial cells lining the mucosal surfaces of intestinal, respiratory and genitourinary tracts, termed as mucosal associated lymphoid tissues (MALT) of the secretory immune system, are equipped to give rise to the synthesis of predominantly IgA or IgE antibodies. If IgA fails to prevent entry of bacteria or parasitic worms into the body, IgE antibodies bound to provide mast cell the next line of defence.
Of lymphocytic cells, the B- and T-lymphocytes are the main cell types that develop from pro to pre and immaturity to maturity in the bone marrow and thymus respectively. The B- and T- lymphocytes, that primarily originate from the bone marrow, migrate to the periphery and tend to aggregate or cluster in the lymphoid tissues. T- lymphocytes colonize the paracortical areas of the lymph nodes and recirculate through blood, i.e. these cells pass through lymphatic network from the periphery to the thoracic duct. Similarly, B- lymphocytes colonize the cortical areas of the lymph nodes as multiple primary follicles and recirculate through blood similar to T- lymphocytes. After primary antigenic stimulation during its passage through the tissue, these cells, for example B- lymphocytes transform into lymphoblasts and the primary follicles differentiate into secondary follicles, also termed as germinal centers. In secondary antigenic stimulation, or booster response, the follicular dendritic cells present antigen or cells complexing through IgGFc or otherwise and expand B-lymphocytes into antibody forming plasma cells and long-lasting memory cells concomitantly.
There are two broad mechanisms of generation of cell surface receptors: one is antigen independent and the other antigen dependent. Antigen independent mechanism of gene reshuffling creates a population of antigen reactive cells or anti-nonself lymphocytes referred to as positive selection. This process also eliminates the population of autoreactive cells or irrelevant anti-self lymphocytes to induce unresponsiveness thus ensuring self tolerance, i.e. tolerance to self or auto-antigens referred to as negative selection. This process takes place in the central lymphoid tissues. Antigen dependent mechanism of genetic reprogramming involves recognition of antigen and stimulation of anti-nonself lymphocytes to induce an immune response and this takes place in the peripheral lymphoid tissues.
In the antigen independent mechanism of gene rearrangement, a small number of antibody genes reshuffle by VDJ recombination and create a great diversity of repertoire of antigen receptors on cell surfaces with specificity to the range of 106 to 109 with an individual. The VDJ recombination, also referred to as somatic recombination, is a process that breaks and rejoins DNA which is involved in the generation of genes for V domains. D genes refer to small genetic elements which recombine with V and J gene to produce a gene for the V domains of antigen receptors. However, anyone lymphocyte can have only one type of receptor that determines specificity, capable of recognizing a small number of antigens, indicating that antigen recognition is not absolute. This process of generation of antigen reactive anti-nonself lymphocytes, representing a large repertoire, occurs in the early stages of the developing lymphocytes.
On entry into the body, the antigen selects one from within the repertoire, which has a complementary configuration and happens to fit it. It means, on maturation and migration to the periphery, the reactive lymphocytes encounter nonself, recognize antigenic substance and select the right determinant to follow the antigen dependent process of genetic reprogramming, involving highly complex sequences of events, deployment of stimulation products and regulation of immune response, particularly in terms of interaction of cells with molecules and consequences of subsequent encounter with antigen.
On stimulation, the antigen causes lymphocyte to transform into blasts and then divide into a number of cells and differentiate into effector and memory lymphocytes, as mentioned earlier, under the influence of lymphokines, giving rise to adaptive immunity or specific acquired response as opposed to the phagocytic cells that give rise to innate immunity or nonspecific protective response singly, in combination or in continuum with acquired immunity. As stimulation progresses, somatic mutation occurs in the antibody genes of activated mature lymphocytes to generate a population of mutant cells with higher affinity for the antigen. In contrast, the immature or irrelevant antiself lymphocytes undergo apoptosis or programmed cell death. This process is referred to as tolerance. Failure of proper editing or screening permit forbidden clones or autoreactive lymphocytes to appear and cause autoimmune diseases.
In this process, autoreactive lymphocytes are eliminated. B-lymphocytes, also called B-cells, produce antibodies; whereas T-lymphocytes, also called T-cells, produce lymphokines, which interact with various immune cells, activate phagocytes to destroy pathogens taken up by the cells, kill target cells infected with virus and cooperate with antigenically activated B-cells for their differentiation into plasma cells. The products of B-cells and T-cells thus regulate immune response to the antigen.20

THE PHYSIOLOGY AND PATHOLOGY OF THE IMMUNE RESPONSE
Immune system is a collection of cells and molecules (Flow chart 1.3.2) which on one hand protect individuals from pathogens as in immunity to infection and on the other cause tissue damage as in hypersensitivity or make them prone to infection as in immunodeficiency.
The molecules of the immune system are constitutive or inducible such as antibodies and cytokines which in an immune response:
- resist infection and give rise to sterile immunity where immunoregulatory mechanism is intact.
- integrate the body as a whole to maintain homeostasis where immunoregulatory mechanism is optimum or normal.
- cause tissue damage, i.e. induce hypersensitivity and give rise to nonsterile immunity, allergy, autoimmunity and transplantation rejection where immunoregulatory mechanisms may go wrong to cause disease and
- fail to resist susceptibility to pathogens causing recurrent or chronic infection where immunoregulatory mechanism is defective, as in immunodeficiency states due to the defect in one or more stages of the development of the cells or synthesis of their molecules.
The immune system may also suffer from suboptimal or decreased immune response as in the two extremes of life such as at infancy, old age, perturbed state of mind or secondary to malnutrition, or disease elsewhere like Cushing's syndrome that release corticosteroids, immunosuppressive therapy with drugs and radiation and viral infection, such as measles and infection with HIV leading to AIDS, which may contribute to the increase in susceptibility to infection or development of autoimmunity and cancer, where immunoregulatory mechanism go wrong or is disturbed.

1.4LANDMARKS IN THE HISTORY OF IMMUNOLOGY
Active immunization with cowpox vaccine led to the discovery of antigen (Edward Jenner, United Kingdom, 1798) and eradication of smallpox, a fatal disease of man, as announced by the World Health Organization in 1979. The Nobel Foundation was set up in 1900 to acknowledge the discovery and commemorate the award-winning contributions in science. The first distribution of the prize took place on 10 December, 1901. Passive immunization with antitoxin in serum proteins similarly led to the discovery of antibody mediated immunotherapy or antitoxic immunity that saved the life of a little girl with terminal diphtheria at Berlin Hospital (1891) and won Emil von Behring (1901) the first Nobel Prize in medicine.
Over the last century, this prize was awarded to twenty others in immunology, a multidisciplinary offshoot of biomedical science. They are in the field of tuberculosis and tuberculin hypersensitivity (Robert Koch, 1905), theory of antibody formation (Paul Ehrlich, 1908), anaphylaxis and histamine (Charles Richet, 1913), complement mediated hemolysis (Jules Bordet, 1919), human blood grouping (Karl Landsteiner, 1930), yellow fever vaccine (Max Theiler, 1951), antihistamine (Daniel Bovet, 1957), acquired immunological tolerance (Macfurlane Burnet and Peter Medawar, 1960), chemical structure of antibody (Gerald Edelman and Rodney Porter, 1972), MHC or HLA (Baruj Benaceraff, Jean Dausset and George Snell 1980), theories of immune function (Neils Jerne, 1984), monoclonal antibody (George Kohler and Cesar Milstein, 1984), genetic basis of antibody diversity (Susumu Tonegawa, 1987) and transplantation of kidney and bone marrow (Joseph Murray and Donell Thomas, 1990).
FURTHER READING
- Benner R, Hijmans W, Haaijman J. Bone marrow: The major source of serum immunoglobulin but still a neglected site of antibody formation. Clinical and Experimental Immunology, 1981;46:1–8.
- Blackwell TK, Alt FW. Immunoglobulin gene rearrangement: mechanism and developmental program of immunoglobulin gene rearrangement in mammals. Annu Rev Genel, 1989;23:605–36.
- Dasgupta A. Immune deviation: Demonstration of split tolerance in vitro by inhibition of macrophage migration. Clinical and Experimental Immunology, 1971;8:173.
- Gowans JL, Knight EJ. Route of re-circulation of lymphocytes in the rat, Proe R Soc Long, B Biol Sci, 1964;159:257–82.
- Lane PJL, Brocker T. Developmental regulation of dendritic cell function, Curr opinion immunol, 1999;11(3):308.
- Marchesi VT, Gowins JL. Migration of lymphocytes through the endothelium of venules in lymph nodes: an electron microscope study, Proc. R Soc Lond, B Biol Sci, 1964;159:283–90.
- Miller JEAP. Thymus: maestro of the immune system, Bioessays, 1994;16:509.
- Przylepa J, Himes C, Kelsoe G. Lymphocyte development and selection in germinal centres. Curr Top Microbial Immund, 1998;229:85–104.
- Ulrich H Von Andrian, Charles R Mackay. T cell function and migration: two sides of the same coin, New Eng J Medicine, 2000;343(14):1020–34.
Books, Reviews and Journals
- Antibodies and Immunity, Basic Books Inc, New York, 1969. Nossal GJV.
- Antigens, lymphoid tissues and the immune response, International series of monograph and treaties, 1971. Nossal GJV, Ada GL.
- Essential Immunology, Oxford: Blackwell Scientific publication, 2001. Roitt IM, Delves P.
- Immunology, Mosby, Harcourt Publishers Limited, 2001. Roitt IM, Brostoff J, Male D.
- Immunology at a glance, Oxford: Blackwell Scientific Publication, 1996. Playfair JH.
- Immunobiology, Churcill livingstone, 2000. Janeway CA, Travers P.
- Modern Immunology, Tata McGraw-Hill 76 and Jaypee Brothers, India, 1993. Dasgupta A.
- Nature, Science, New England Journal of Medicine, Journal of Experimental Medicine, Immunological Reviews Immunology Today, Immunologist, Current opinion in immunology, Clinical and experimental immunology, Immunology, Journal of Immunology, Clinical and experimental immunology, Cellular immunology, Infection and immunity, European journal of immunology, Parasite immunology, Journal of allergy and clinical immunology, Tissue antigens, Immunogenetics.
- Clonal Selection Theory of Acquired Immunity, Cambridge University Press, London, 1959. Burnet FM.
- Lymphocyte-a disgraceful gap in medical knowledge, Immunology Today 1996; 17, 288-291. Gowans JL.
- Specificity of Serological Reactions, Harvard University Press, Boston, 1964. Landsteiner K.
- Immunity in the Infectious Disease, Macmillan Press, New York, 1905. Metchnikoff E.
- History of Immunology, Academic Press, London. Silverstein AM.
- Molecular Biology of the Cell, Garland Publishing, New York, 1994. Alberts B, Bray D, Lewis J, Raff M, Roberts K, Watson JD.