


Over the years the ideas about cell structure have changed. Biologists in the past saw cells as simple membranous sacs containing fluid and a few floating particles. Today's biologists know that cells are infinitely more complex than this.
Human body has cells of different types, sizes, and shapes. The concept of a “generalized cell” for descriptive purpose includes features from all cell types. A cell consists of three main parts: the cell membrane, the nucleus, and between the two, the cytoplasm. Within the cytoplasm lie intricate arrangements of fine fibers and hundreds or even thousands of minute but distinct structures called organelles.
TYPES OF CELLS
Depending on the fundamental structure there are two primary types of cells; prokaryotic cells and eukaryotic cells. Prokaryotic cells have no true nucleus while eukaryotic cells have a true nucleus. There are many distinctions between these two different types of cells as summarized in Table 1.1.

Prokaryotes are organisms made up of cells that lack a cell nucleus or any membrane-encased organelles. Most prokaryotes are unicellular, but there are a few that are made of collections of cells (multicellular).
Eukaryotes are organisms made up of cells that possess a membrane-bound nucleus as well as membrane-bound organelles (Fig. 1.1).
Prokaryotic Cells are Small and Simple but Fast Multiplying
Size: Mycoplasma is 0.1 to 1 micron in diameter; other bacteria are 1 to 10 micron in diameter. A few much larger exceptions are known. Small cells such as bacteria divide faster (once in 20 min). Have no nucleus, hence DNA is less protected and mutates faster. Human body is not made of prokaryotic cells, but gut is inhabited by approximately 1.3 kg of bacteria. Bacteria inhabiting our gut are small in size and outnumber our body cells by a factor of 10:1. Intestinal bacteria are believed to be beneficial, because they produce vitamins and stimulate the immune system.
Eukaryotic Cells are Large and Slow Multiplying but Versatile
Size: typically 10 to 100 micron in diameter and are much larger than prokaryotes. Cell division is very slow (once in 20 hours). Have 5nucleus, hence DNA is better protected and mutation rate is low. Organelles are present, which allow many activities to take place within the same cell but in different compartments. Organelles can be isolated for study by centrifuge techniques.
There are about 250 Types of Specialized Cells in the Body
Cells specialize by turning their genes on and off, and by structural modifications.
Few examples of specialized cells are:
Red blood cells (matured) have lost their nuclei and mitochondria, and are specialized for transport of gases in blood. These cells are loaded with hemoglobin (O2 carrying protein) specialized for carrying O2 to the tissues.
Nerve cells have long axons measuring a meter or more in length and are specialized for transmitting electrical impulses, they have specialized Na+ and K+ channels for generating electricity. These cells have a single nucleus in the cell body with special axonal transport mechanism to deliver proteins made in the cell body to the ends of the cell.
Muscle cells have special contractile proteins actin and myosin arranged in a sarcomere, which is specialized for producing force by contraction. These are very long cells often attached to two bones. Muscle cells are formed by fusion of many smaller cells, so they have many nuclei.
Insulin-secreting cells (beta cells of pancreas) have large amounts of rough endoplasmic reticulum needed for insulin synthesis. Gene for making the insulin hormone is turned on in beta cells of Langerhans.
What are the Biological Advantages of Having Cell Organelles!
Some of the Organelles Found in Eukaryotic Cells are the Result of Endosymbiosis during Evolution
Cells often ingest other cells and digest them for food. Sometimes the ingested cell is not digested, but the two cells learn to live together for mutual benefit (endosymbiosis). Mitochondria and plant chloroplasts are believed to have originated in this way. These organelles have their own DNA and plasma membrane made of lipid bi-layer.
Cell Organelles are found within the Cytosol
Cytosol is the liquid matrix of the cell, which is mostly water that contains cell organelles (except nucleus), salts, dissolved molecules, and enzymes. Glycolysis (anaerobic energy metabolism) takes place in the cytoplasm.6
Cell Membrane Separates the Cytoplasm from its Surrounding
Cell membrane is a lipid bi-layer made of phospholipids and proteins. It forms a barrier for the movement of molecules in and out of the cell; hydrophobic molecules pass through it more readily than hydrophilic ones. Specialized transport mechanisms allow selective movement of materials across the membrane.
Nucleus Contains the Molecule of Heredity (DNA)
The nucleus is the control center of the cell that determines how the cell will function, as well as the basic structure of that cell. It has a porous outer membrane called nuclear envelop that encase the fluid nucleoplasm within which are found the threads of chromatin (DNA)—the genetic material of the cell, and the nucleolus. There are many proteins associated with DNA, which are involved in DNA repair and turning the genes on and off. The nucleolus is within the nucleus and is the site of ribosome formation. It has copious amounts of RNA and proteins. The nucleolus also contains nucleolar organizers which are parts of chromosomes with the genes for ribosome synthesis.
The nuclear envelop is a double membrane that separates the contents of the nucleus from the cytoplasm. The envelope helps to maintain the shape of the nucleus and assists in regulating the flow of molecules (including RNA) into and out of the nucleus through nuclear pores. When a cell is “resting” (not dividing), the chromosomes are organized into long entangled structures called chromatin and not into individual chromosomes as we typically think of them.
Most cells contain single nucleus, but a few have more. Examples are: some liver cells have multiple nuclei (polyploidy); muscle cells are very long and have hundreds of nuclei. Mature red blood cells have lost their nucleus.
Centrioles Organize the Mitotic Spindle for Cell Division
Centrioles are a pair of small structures found in the centrosome near the nucleus; its structure is similar to that of cilia made of 9 triplet tubules. In animal cells, centrioles divide before cell division and help to organize the mitotic spindle made of tubulin.
Mitochondria are the Powerhouses of the Cell
Mitochondria are the sites of cellular respiration which ultimately generates fuel for the cell's activities. Hence are called the “power houses” of a eukaryotic cell. They convert energy into forms that are usable by the cell
Mitochondria are sausage shaped organelles that are 0.5 to 10 µm long. They are double membranous bags of proteins involved in developing chemoelectrical gradient to generate cellular energy. The outer membrane is smooth and contains porin (transmembrane 7protein) that allows passage of molecules up to 5,000 Dalton. The inner membrane is thrown into folds to form cristae and is selectively permeable, which generates proton gradient to produce heat that in turn generates ATP via oxidative phosphorylation (Fig. 1.2). The folds enhance the “productivity” of cellular respiration by increasing the available surface area. The intercristae space or matrix (space bounded by inner membrane), contain enzymes for Krebs cycle, lipid and protein synthesis, mitochondrial DNA, RNA and ribosomes. The intermembrane space is the narrow part between the two membranes.
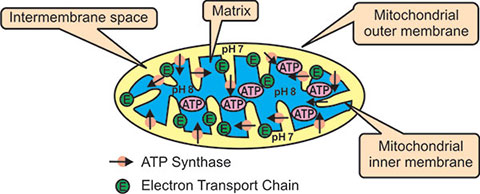
Mitochondria are semiautonomous in that they can divide and grow to make more of themselves. They also have their own DNA and ribosomes.
Mitochondrial matrix contains dehydrogenases and mitochondrial DNA that is always inherited from the mother. Thus transmission of diseases of energy metabolism is always from the mother.
Krebs cycle reactions take place in the internal matrix (blue). NADH and FADH2 (produced by glycolysis and the Krebs cycle) deliver their hydrogens and electrons to the electron transport chain (ETC). The ETC pumps hydrogen ions into the intermembrane space (yellow); this sets up a pH gradient- pH 8 in the matrix and pH 7 in the intermembrane space. Hydrogen ions flow through a channel in the enzyme ATP synthase from the intermembrane space to the matrix (see arrows). This causes a protein shaft in the ATP synthase to rotate and generates ATP in the matrix.
The Peroxisomes (Microbodies)
Peroxisomes are membrane bound spherical vesicles that are abundant in parenchyma of liver and kidney. They measure 0.5 to 1.2 µm. They contain oxidase and catalase that remove hydrogen 8peroxide, and also use hydrogen peroxide to peroxidize organic substances via catalase, beta oxidation of lipids, and detoxification of organic compounds such as ethanol. The enzymes of peroxisome are synthesized in the cytosol and sorted to peroxisome via a signal amino acid sequence of -Ser-Lys-Leu near carboxyl terminal and receptor on peroxisome for this sequence.
Proteins are made on the Ribosomes
Ribosomes are 20–30 nanometer particles composed of ribosomal RNA and protein, and are made in the nucleolus. Each unit of ribosome has two subunits. On electron microscopy they appear as beads on a string; beads are the translational protein machinery and string is the mRNA. On light microscopy ribosomes are seen as basophilic bodies, Nissl's substance in neurons, and ergastoplasm in glandular cells. They are involved in protein synthesis either as free polyribosomes in the cytosol or as bound polyribosomes on the endoplasmic reticulum (rough ER). Free polyribosomes synthesize intracellular proteins, and the bound (rER) polyribosomes synthesize proteins destined for the extracellular space or within the membrane. Rough ER is prominent in cells that are secreting hormones and enzymes (e.g. pancreatic cells).
The Endoplasmic Reticulum
Endoplasmic reticulum (ER) is an extensive network of membranes composed of membrane bound enzymes, receptors, and transporters. They appear as elongated, flattened, rounded or tubular vesicles that have either a smooth or rough surface. The two regions of ER differ in both structure and function. The regions of ER to which ribosomes are attached on the cytoplasmic side of the ER membrane are called rough ER because it gives a rough appearance. The other region is called smooth ER because it lacks attached ribosomes. Typically, the smooth ER is a tubule network, and the rough ER is a series of flattened sacs (Fig. 1.1). The two regions are usually interconnected and the proteins and membranes made by the rough ER move into the smooth ER to be transferred to other locations.
The smooth ER has a wide range of functions including carbohydrate and lipid synthesis. It serves as a transitional area for vesicles that transport ER products to various destinations. In liver cells the smooth ER produces enzymes that help to detoxify certain 9compounds and glycogen metabolism. In muscles it stores calcium that assists in the contraction of muscle cells. In adrenal cortex and gonad cells it synthesizes steroid hormones. It is also involved in lipid metabolism and membrane formation.
The rough endoplasmic reticulum is involved in protein synthesis, protein sorting, and initial post-translational modification such as glycosylation, sulfation, phosphorylation, and hydroxylation of proteins, and also the synthesis of phospholipids. Protein sorting involves several factors and a sequence of events.
Golgi Apparatus Finishes, Sorts, and Targets Proteins
In eukaryotic cell, the Golgi complex or Golgi apparatus is responsible for manufacturing, warehousing and shipping certain cellular products, particularly those from the endoplasmic reticulum (ER). It is hence called the “manufacturing and shipping center”. Depending on the type of cell, there can be just a few complexes or there can be hundreds. Cells that specialize in secreting various substances typically have a high number of Golgi complexes.
A Golgi complex is a membrane delimited organelle composed of flat stacked sacs containing various enzymes. Each stacked grouping has a smooth membrane that separates the insides from the cell's cytosol.
The Golgi complex has polarity. Membranes at one end of the stack differ in both composition and in thickness from the other end. One end that acts as the “receiving” (Cis face) department is closely associated with ER. The other end is the “shipping” (Trans face) department, which produces vesicles that “ship” materials to other sites.
The Golgi complex modifies many products including proteins and phospholipids. Post-translational modifications of proteins include, terminal glycosylation, sulfation, phosphorylation, and limited proteolysis. The Golgi apparatus can also manufacture certain types of macromolecules. One such example is hyaluronic acid which is a form of cellular glue.
Golgi complex is also involved in sorting and addressing/targeting (sending) proteins to their specific locations such as: transport to lysosomes, incorporating into the cell membrane, and transport into secretory vesicles.
Some of the intracellular addressing systems are as follows:
- Proteins with the signal sequence of -Lys-Asp-Glu-Leu-COOH are retained in ER/Golgi complex.
- Proteins that undergo post-translational modification such as phosphorylation of mannose at the C6 are destined to lysosomes, and are segregated into regions that pinch off and form compartments with mature lysosomal enzymes.
- Proteins without address enter default pathway into secretory vesicles for exocytosis.
Lysosomes Digest Materials within the Cell
Lysosomes are membranous sacs of enzymes. These enzymes are typically hydrolytic and can digest cellular macromolecules. They are made by the endoplasmic reticulum and Golgi complex. Lysosomes are formed by budding from the Golgi complex. Lysosomes contain various hydrolytic enzymes (acid hydrolases active at pH 4.5) that are capable of digesting nucleic acids, polysaccharides, fats and proteins. The inside of a lysosome is acidic. Since the enzymes work best in an acidic environment, if a lysosome's integrity is compromised, the enzymes would not be very harmful in the neutral cytosol.
Lysosomes have various roles. They fuse with late endosomes to facilitate degradation of endosome contents. They are active in recycling the cell's organic material and in the intracellular digestion of macromolecules. In addition, in many organisms, lysosomes are involved in programmed cell death.
Cell Shape is Supported and Determined by Cytoskeleton
Tubules are imbedded in the cytosol to form a meshwork of fibers that give shape to the cell. These tubules help the transport of structures within the cell (example: movement of chromosomes during cell division) and are also involved in movement of the whole cell. Also attaches to intercellular connections linking cells together.
The cytoskeleton is composed of at least three different types of fibers: microtubules, microfilaments and intermediate filaments. These 11types are distinguished by their size. The microtubules are the thickest and microfilaments are the thinnest.
- Microtubules (25 nm in diameter) are hollow rods functioning primarily to help support and shape the cell and as “routes” along which organelles can move. Microtubules are typically found in all eukaryotic cells.
- Microfilaments or actin filaments (7 nm in diameter) are solid rods and are active in muscle contraction. Microfilaments are particularly prevalent in muscle cells but, are typically found in all eukaryotic cells.
- Intermediate filaments (8 to 12 nm in diameter) can be abundant in many cells. They provide support for microfilaments and microtubules by holding them in place.
In addition to providing support for the cell, the cytoskeleton is also involved in cellular motility, movement of vesicles within a cell, and assists in the formation of food vacuoles.

Cilia and Flagella Help Cell Movement
Eukaryotic cilia and flagella are whip-like projections from the cell. Both have a core, composed of microtubules connected to the plasma membrane arranged in what is known as a 9 + 2 pattern (Fig.1.3). The pattern is so named because a ring of nine microtubule “doubles” has in its center two singular microtubules. This organization allows the sliding of the microtubule doubles against one another to “bend” the cilia or flagella. Both cilia and flagella are found in various types of cells. For instance, the sperm of many animals have flagella. Cilia can be found in cells lining respiratory and female reproductive tracts.
Differences between Cilia and Flagella
The protrusions that are short and numerous are termed cilia. If they are longer and less numerous (usually only one or two) are termed flagella. Movement of flagellum is quasi-sinusoidal, whereas the movement of cilia is whip-like.
Flagellated cells usually have only 1 or 2 flagella measuring 50 to 200 microns in length. Example: sperms.
Ciliated cells usually have hundreds of cilia that measure 5 to 10 microns in length. Examples are: cells lining respiratory tract that helps expel mucus secretions, cells lining fallopian tube that help to move egg cells, cells lining spinal canal that help move cerebrospinal fluid.
Microvilli Increase the Surface Area of Cells
Projections of cell surface form the brush borders of cells are sometimes confused with cilia, but they are much shorter (1 micron length) and with a different structure. Projections are supported by cytoskeleton filaments mostly the protein actin. Such projections increase the cell surface for faster absorption and secretion of materials.
Example of absorptive cells with microvilli is intestinal epithelium cells. Examples of secretory cells with microvilli are choroid plexus cells of brain that secrete cerebrospinal fluid.
Specialized microvilli called stereo cilia (misnamed) are found on the surface of the hair cells of the inner ear. The stereo cilia respond to sound vibrations and are involved in hearing.
PLASMA MEMBRANE

Within the phospholipid bilayer, cholesterol, glycolipids, proteins and glycoproteins are interspersed (Fig.1.4).
Phospholipids are amphipathic; fatty acid tails are attached to hydrophilic head group, examples are phosphotidylserine, phosphotydylcholine, phosphotidylethanolamine, and sphingomyelin. Presence of cholesterol in the lipid bilayer improves membrane rigidity and prevents crystallization at low temperatures. Gangliosides, an example of glycolipid are found on extracellular surfaces of membranes. The differences in the carbohydrate moieties of glycolipids are used for blood typing.
Proteins that span the entire thickness of the membrane are called integral proteins. They function as specific ion channels (Na/K ATPase) or selective transporters (glucose transporters). The proteins that are attached to only one side of the membrane are called peripheral proteins (examples is spectrin). The proteins exposed to the extra-cellular side of the membrane have carbohydrates chains attached covalently.
The branched polymers of carbohydrates, usually found on the extracellular surface of membranes include galactose, N-acetylgalactosamine, N-acetylglucosamine, fucose, mannose, and sialic acid. They are covalently linked to membrane proteins and lipids. The composition of these carbohydrate polymers are specific to given cell type that are recognized by specific proteins. For example, carbohydrate moieties of selectins on endothelial cell are recognized by peripheral proteins on neutrophils.
Features of Plasma Membrane
Plasma membrane is about 10 nanometers in thickness. The lipid bilayer is impermeable to most polar substances (ions, glucose, etc.) 14but permit passage of water. Lipid bilayers are 2-D fluids in which the sideway or lateral movements of molecules within the leaflet is very fast, but the transverse movement across to apposite leaflet is slow. Membrane fluidity is a feature of importance for biological membrane. The membrane fluidity depends on temperature. At higher temperature membrane is fluid since acyl tails are randomly arranged. At lower temperature it is solid since acyl tails are organized. Phase transition from a liquid to gel and gel to solid with a change in temperature, depends on the acyl chain length (with increasing acyl chain length Van der Wall's interaction increases, and more energy is needed to pull apart acyl chains, hence solid even at higher temperature), degree of saturation (with increasing number of double bonds steric hindrance prevent close packing of acyl chains and Van der Wall's interaction decreases, hence liquid even at lower temperature) of acyl chains and amount of cholesterol (an increased cholesterol content inhibits crystallization of acyl chains by fitting between them and prevent gel formation even at colder temperatures) in membrane.
In 1972, Singer and Nicholson postulated that biological membranes resemble proteins floating in a 2D lipid sea (Fluid Mosaic Model of membrane structure). Biological membranes act as permeability barrier and solvent for integral membrane proteins. These proteins could be peripheral, integral or transmembrane. Integral proteins are embedded through or within membrane tightly bound by hydrophobic forces, peripheral are attached to membrane. Membrane proteins are amphiphilic, they have both polar and nonpolar regions hence are asymmetrically arranged, i.e. attached to one side, but located within the membrane. Both proteins and lipids are glycosylated (e.g. gangliosides). Lipids are asymmetrically distributed; most common externally exposed lipids are sphingomyelin and phosphotidylcholine. Lipids exposed to the interior are phosphotidylethanolamine. Integral proteins and lipids rapidly diffuse within the plane of the membrane. Lateral mobility of a lipid is fast but Flip-flop of lipids (from one half of a bilayer to the other) is very slow. Flip-flop would require the polar head-group of a lipid to traverse the hydrophobic core of the membrane. Flippase enzymes catalyze flip-flop in membranes where lipid synthesis occurs. Otherwise, flip-flop of lipids is rare. The two leaflets of a membrane bilayer tend to differ in their lipid composition. Flip-flop of integral proteins does not occur. All copies of a given type of integral protein have the same orientation relative to the two sides of the bilayer membrane.
Defective Cell Organelles are Responsible for Some Diseases
- Tay-Sachs is a lysosomal storage disease in which gangliosides accumulate in lysosomes since they lack hydrolases that are responsible for its breaking down.
- Abnormal mitochondria with defective aerobic metabolism can lead to accumulation of lactic acid that causes lactic acidosis.
- Hereditary elliptocytosis of erythrocytes is an uncommon disease due to defective anchore protein. α-spectrin found in erythrocyte cell membrane resulting in oval shape of RBC, and hemolysis.
CELL DIVISION
Reasons for Cell Division
Cell division is required for: (a) growth, (b) repair and replacement of damaged parts, and (c) reproduction of the species.
In Cell Division Copies of the DNA must be sent to both New Cells
Since the instructions for making cell parts are encoded in the DNA, each new cell must get a complete set of the DNA molecules. This necessitates the DNA be copied (replicated or duplicated) before the cell undergo division.
Genetic Blueprints for Cells are Organized into Chromosomes
The plans for making cells are coded in DNA, which is organized into giant molecules called chromosomes. Each chromosome is a single DNA molecule containing many genes. Each gene gives the directions for making one protein. In humans, each chromosome has approximately 2000 genes.
Chromosomes have Distinct Parts
Centromere is the part that holds duplicated chromosomes together before they are separated during mitosis. Kinetochore proteins bind to centromere of chromosome to form spindle during mitosis. Telomeres are the ends of chromosomes important in cell aging.
DNA in chromosomes is associated with proteins. These proteins strengthen DNA fiber, help package chromosomes when they condense, and control activity of genes.
Human's Cells have 23 Pairs of Chromosomes (Diploid = Pair of each Chromosome = 46 total)
The members of a chromosome pair are called homologues. One of each pair comes from mother, the other from father. Human reproductive cells (sperms and eggs) have 23 single chromosomes (Haploid = single copy of each chromosome = 23 total).
Genetic Instructions are Organized into Genes
A section of DNA, which codes for a protein is called a gene (One gene, one enzyme). We have approximately 50,000 genes (approx. 2000 per chromosome). Most of DNA in chromosome (about 95%) is “junk” DNA whose function not known.16

Before a Cell can divide it must Duplicate its Chromosomes
To make a new cell the old cell must duplicate all its parts. Duplication of all cellular organelles takes place in interphase, but DNA (chromosomes) is duplicated in the ‘S’ sub-phase. Entire chromosome is duplicated at the same time. The duplicated chromosome remains attached to the original chromosome by its centromere. The original chromosome and its duplicated partner are called sister chromatids (Fig. 1.5).
During Duplication the DNA Strands Separate (Unzip)
DNA is a double helix spiral with the two strands held together by hydrogen bonds. During replication the two strands come apart and each acts as a template to form a new strand. The coming apart (unzipping) is made possible because the strands are held together by hydrogen bonds.
Chromosomes must be Tightly Packaged for Division
DNA must be tightly packaged for division otherwise it would tangle. This is made possible by histones and other proteins on which DNA is wound up to enable strands get 10,000 times shorter and much thicker (called condensation). Condensation occurs during prophase and they become visible under microscope.
Mitosis is used for Growth and Repair
Object of mitosis is to produce two identical cells with same number of chromosomes. This is required for growth, repair and reproduction (in single-cell organisms). During mitosis cell divides only once after DNA duplicates, giving each cell the original number of chromosomes.
Some cells such as those lining the gut and skin, and white blood cells must be repaired often; whose cell lifespan is only a few days.
Others such as nerve and muscle cells do not divide at all after birth.
Red blood cells have an intermediate lifespan of about 120 days.
General Scheme of Mitosis
The DNA duplicates to form two sister chromatids, then chromosomes attach to spindle and separate (Fig. 1.6).
Meiosis is used for Sexual Reproduction
Object of meiosis is to reduce the number of chromosomes to single copy (haploid) of each, for making sperm and eggs that are haploid. When a sperm fertilizes an egg to form a zygote the diploid number of chromosomes is restored (23 + 23 = 46). In meiosis, cells divide twice after a single DNA duplication.

FIGURE 1.6: General scheme of mitosis (2D = DNA content of diploid cell; 4D = amount after duplication)
General Scheme of Meiosis
DNA duplicates to form 2 sister chromatids, then chromosomes attach to spindle and separate. First division separates homologue chromatids, and the second division separates sister chromatids, which results in 4 haploid cells each with half the original number of chromosomes (Fig. 1.7).

FIGURE 1.7: General scheme of meiosis (2D = amount of DNA in diploid cell; 4D = amount after duplication; 1D = amount of DNA in haploid cells (sperm and eggs)
Spindle Apparatus Separates Chromosomes both in Mitosis and Meiosis
Spindle is formed of microtubule that attach to chromosomes at centromere. Separation of chromosomes requires energy (ATP).
Five Stages of Cell Cycle
Interphase is the longest phase during which essentially the cell must duplicate all its organelles. During this phase proteins synthesized, centrioles duplicated, and chromosomes replicated (sister chromatids) but sister chromatids remain attached at centromere until anaphase.
Prophase is the phase during which chromosomes condense and become visible, centrosomes move to opposite ends of cell to form spindle, and nuclear membrane dissolves.
Metaphase: Chromosomes attach to spindle fibers at their centromeres, and line up in center of spindle apparatus.
Anaphase: Centromeres split to free the sister chromatids, and chromosomes move toward centrosomes to opposite poles of cell.
Telophase/Cytokinesis: Cell cleaves to form two cells (cytokinesis), and the nuclear membrane reforms. The last four phases are called mitosis (Fig. 1.8).
Mitosis = Prophase + Metaphase + Anaphase + Telophase (PMAT)

Cancer is Uncontrolled Mitosis
Mitosis must be controlled otherwise growth will occur without limit (cancer). Special proteins produced by oncogenes control cell growth. Mutations in such oncogenes can cause cancer.
CELL DEATH
The irreversible injury to cell, results in cell death that usually takes the form of necrosis. The structural changes that accompany necrosis result from two processes:
- Enzymatic digestion of the cell by its own hydrolytic lysosomal enzymes (sometimes called liquefaction necrosis).
- Denaturation and precipitation of cellular proteins (coagulation necrosis).
Liquefaction necrosis occurs in some bacterial infections (e.g. Staphylococcus) and ischemic injury to brain tissue. Coagulation necrosis is a common manifestation of hypoxic cell injury (e.g. myocardial infarction). Tuberculosis infections produce a combination of liquefaction and coagulation necrosis characterized by a collection of soft, whitish-gray debris resembling clumped cheese (caseous necrosis). Necrosis is accompanied by inflammation and secondary injury to surrounding normal tissues.
Necrosis and Apoptosis
Necrosis should be distinguished from apoptosis; genetically programmed cell death. In apoptosis, there is an orderly disassembly of cellular proteins and DNA with minimal disruption to normal tissue. Apoptosis is a normal physiologic process designed to eliminate unwanted, functionally abnormal, or senescent cells. It plays an important role in the developing embryo (fashioning out fingers from limb buds), certain hormone-dependent tissues (denudation of endometrium during menstruation), and in aging. However, in some instances, apoptosis may be a pathologic process induced by cell injury (e.g. viral infection, radiation injury, etc.).
Apoptosis is programmed cell death: physiological process involved in development and other processes:
- DNA fragmentation at specific sites
- cell volume decrease
- cell membrane blebbing
- formation of apoptotic bodies.
Table 1.2 summarizes some of the important differences between necrosis and apoptosis. Necrosis is a “messy” cell death, while apoptosis is a more orderly process.19
|
FUNCTIONS OF CELL MEMBRANE
Cell Membranes are Primarily Phospholipid Bi-layers (Two Layers)
Phospholipid bi-layer forms a barrier to passage of molecules in and out of cell. In addition, presence of cholesterol in the lipid bi-layer stabilizes cell membranes. In Figure 1.9 note that the hydrophobic tails of the phospholipids (fatty acids) are packed together in the center of the bi-layer that keeps them out of water.

Membranes also Contain Proteins
Proteins that penetrate the membrane have hydrophobic sections of about 25 amino acids long (Fig. 1.10). Membrane proteins functions as:
- Receptors for hormones.
- Ion channels.
- Adhesion molecules for holding cells to extracellular matrix.
- Cell recognition antigens.
All Molecules move Continuously by Simple Diffusion
Heat energy causes molecules to move randomly. If the concentration of molecules is different at two regions, diffusion will cause molecules to move from a region of high concentration to one of low concentration. Higher the concentration gradient more rapid is the net diffusion. Diffusion tries to even out the concentrations so that they are equal everywhere.
Simple Diffusion Across a Membrane is called Permeability
Diffusion is most efficient over short distances about the diameters of cells. This type of penetration does not require energy from ATP. Hydrophobic substances have a high permeability through lipid bi-layer membranes, since they cross membranes faster than those that are hydrophilic (Fig.1.11). Many chemicals used in biological system are deliberately made hydrophobic to increase their rate of penetration into cells. Examples: many drugs, pesticides such as DDT.

Osmosis Moves Water Across Biological Membranes
Osmosis is movement of water from low osmotic pressure (dilute solution) to high osmotic pressure (concentrated solution). Except for blood flow, almost all water movement in body is by osmosis. It is useful to think of a dilute solution as having a high water concentration and a concentrated solution as having a lower water concentration. Then the water flow goes from high water to low water concentration. 21In Figure 1.12, the blue dots represent the solute (the higher the solute concentration the lower the water concentration).
Osmosis is Passive as it does not Require ATP Energy
Osmotic flow through most biological membranes is not by simple diffusion, it is by bulk flow and is similar to the flow caused by a pressure gradient. The kidney is an osmotic machine it adjusts body water volume by osmosis. Medical problems involving osmosis are pulmonary edema, childhood diarrhea, cholera, and inflammation of tissues.

CLINICAL CORRELATE
Cholera is caused by Osmotic Imbalances, which can be treated using the Principle of Osmosis

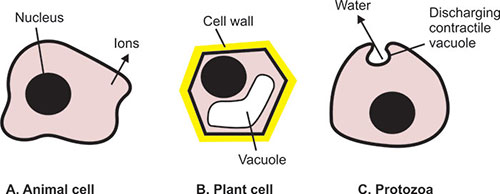
Cells Swell in Hypotonic Solutions and Shrink in Hypertonic Solutions
Osmosis often produces significant volume changes, causing swelling or shrinking of cells. If the external solution balances the osmotic pressure of the cytoplasm it is said to be isotonic. If the external solution is more dilute than the cytoplasm it is hypotonic, and if the external solution is more concentrated it is hypertonic.
- The plant cell is saved from swelling and bursting by its tough wall.
- The protozoa avoids swelling by periodically ejecting water out of the cell.
In Facilitated Diffusion Special Proteins Help to Move Substances Across Membranes
Protein transport molecules help to carry many substances across membranes.
In Figure 1.16 an extracellular molecule (glucose) is shown binding to the transport protein; the transport protein then rotates and releases the molecule into the inside of cell.
Examples: There are five different glucose transporters (GLUT) proteins, and two cotransports (Na+ and glucose cotransporters that are used for secondary active transport).
There are eight different types of aquaporins (water channels).

Properties of Facilitated Diffusion
Facilitated diffusion does not require ATP energy, but cannot cause net transport of molecules from low to high concentration, which would require energy input. Transport rate reaches a maximum when all transporter proteins are being used (saturation). These systems are very specific: allows cell to select substances to be taken up and are sensitive to inhibitors that react with protein side chains. System is of ancient origin: found in all organisms and is extremely important in physiology: about one-third of our basal metabolism is used in active transport of various substances.
Active Transport Uses Energy to Pump Molecules against Concentration Gradient
Pumps are proteins that use energy to carry substances across the cell membrane and can transport substances from a low concentration 24to a high concentration (“uphill” transport) and require energy in the form of ATP. Examples: the Na+/K+ pump, the Ca++ pump, etc.
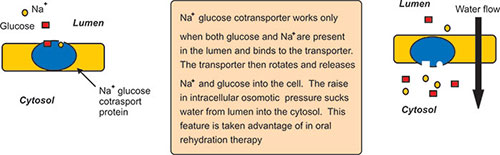
Many Molecules Enter Cells by Secondary Active Transport
Secondary active transport is a combination of active transport and facilitated diffusion. Sodium gradient is produced by the Na+ pump (active transport). The concentration gradient thus produced is used to produce secondary transport of sugars and amino acids (facilitated diffusion). Some sugar and amino acid transporters must bind Na+ as well as the sugar or amino acid (coupled transport) simultaneously. Both Na+ and the organic molecule must be present at the same time and on the same side of the membrane, since there is more Na+ outside the cell, sugars and amino acids get transported mainly from the outside to the inside. The sugar and amino acid transporters do not use ATP directly, but ATP is required to set up the Na+ gradient.
Examples: Glucose transport across the cell wall lining the gut. Cells regulate permeability by adding and removing membrane transport proteins. If a molecule is moved across the cell membrane by a protein transporter or pump; adding more transporters or pumps will increase the transport rate, whereas removing transporters or pumps will decrease the transport rate. Often transport proteins are stored in vesicles until they are needed (Fig. 1.17).

The transporters are down regulated, back into vesicles, when they are not needed. In other cases, new transporters must be synthesized when needed. The body uses hormones to regulate membrane transport in this way. Examples are shown in Table 1.3.25
|
Endocytosis can bring Macromolecules into the Cell
In endocytosis (Fig.1.18), the cell membrane bends inward (invaginates), forming a vesicle containing extracellular fluid, which can bring in large molecules such as proteins. The macromolecules are usually digested by lysosomes.

Cells Do not end at their Outer Membrane; They Possess an Extracellular Matrix (Fig. 1.19)
- Animal cells do not have walls, but do have extracellular matrix; meshwork of macromolecules outside plasma membrane. Consists mainly of glycoproteins (proteins with oligosaccharide chains), especially collagen.

Cells are Joined by a Variety of Intracellular Junctions
In multicellular organisms, adjacent cells are held together by several types of specialized junctions (Fig.1.19).
- Tight junctions: Specialized “belts” that bind two cells tightly to each other, prevent fluid from leaking into intracellular space.
- Desmosomes: Intercellular “rivets” that create tight bonds between cells, but allow fluids to pass through intracellular spaces.
- Gap junctions: Formed by two connecting protein rings embedded in cell membrane of adjacent cells. Allows passage of water and small solutes, but not macromolecules (proteins, nucleic acids).
