

section I
Clinical Strabismus
(Brazil)
Binocular vision is the integration of the sensation produced by the light stimulus in each eye into a unified perception. It is an acquired, conditioned reflex, which has some requisites for its development:
- There must be proper fixation with each eye. The muscles controlling the movements of each eye must function normally and turn both eyes in such a manner that the object of regard is fixated by corresponding retinal areas, e.g. the two foveas.
- The visual fields of the two eyes must overlap to a large extent.
- Approximately similar images must be formed on each retina. The images, therefore, must be approxi-mately of the same size, shape, color, and intensity.
- The retinas must possess physiologically corres-ponding points, i.e. retinal receptors which are so related that they have a common visual direction.
- The eyes must be coordinated by the reflex activities which produce the fusional movements at all times so that retinal receptors which have a common visual direction will receive the same image at all times.
Based on the classic teaching of Claude Worth around the turn of the century, binocular vision has traditionally been broken down into three levels or grades:
- Simultaneous binocular perception,
- Fusion, and
- Stereopsis.
SIMULTANEOUS BINOCULAR PERCEPTION
Simultaneous binocular perception in the context of Worth's scheme refers to the subjective visual superimposition of dissimilar objects viewed by the two eyes. An example of this lowest grade of binocularity is the ability to perceive a lion in a cage (Fig. 1) when viewing the cage with one eye and the lion with the other through a synoptophore (Fig. 2).
Simultaneous perception of differing binocular images is limited by the phenomenon of rivalry, which may exclude from visual awareness part of the input from one fovea or the other when the two differ sufficiently.
If a person looks into a stereoscope at two dissimilar targets with overlapping nonfusible contours, first one contour, then the other will be seen, or mosaics of one and the other, but not both contours simultaneously. In Figure 3, taken from Panum, each eye sees a set of oblique lines, one going from above right to below left, seen by the left eye (Fig. 3A), and another set going from above left to below right, seen by the right eye (Fig. 3B). When observed in a stereoscope, these lines are not seen as crossing lines but as a changing pattern of patches of oblique lines going in one or the other direction (Fig. 3C).
It is of interest that it takes a certain buildup of time (150 m) before dissimilar visual input to the eyes causes binocular rivalry. Dichoptic stimuli were perceived as “fused” when presented for shorter periods.
The phenomenon of retinal rivalry is basic to binocular vision and may be explained as follows. Simultaneous excitation of corresponding retinal areas by dissimilar stimuli does not permit fusion; but since such excitations are localized in the same visual direction and since two objects localized in the same place give rise to conflict and confusion, one or the other is temporarily suppressed. 2Which of the two is suppressed more depends on the greater or lesser dominance of one eye rather than on the attention value of the visual object seen by each eye. In other words, it is the eye and not the stimulus that competes for dominance under a wide range of conditions. Stimulus rivalry occurs only within a limited range of spatial and temporal parameters.
It is at once clear that rivalry phenomena, or rather their absence, must in some fashion be related to what is known as suppression in strabismic patients. Constant foveal suppression of one eye with cessation of rivalry leads to complete sensory dominance of the other eye, which is a major obstacle to binocular vision. Return of retinal rivalry is a requisite for re-establishment of binocular vision.
The retinal rivalry phenomenon has been explained in neurophysiologic terms by the presence of separate channels for the right and left eyes that compete for access to the visual cortex. A third binocular channel is activated only by fusible input. Because of this competition and the inhibition elicited, only fragments of the image seen by each eye are transmitted to the striate cortex in the case of nonfusible binocular input. Competitive interaction occurs not only in the primary visual cortex but continues at several afferent levels of the visual pathway, well after the inputs to the two eyes have converged.
In the clinical setting, normal rivalry can be evaluated by looking closely at the white spot in the worth four-dot display through the red-green viewing glasses (Fig. 4). A shifting combination of red and green areas is seen within the spot, rather than the uniform (yellow) color that would result if the red and green images were physically superimposed on the retina.
When rivalry is in effect, input from the involved foveal region is excluded from conscious perception regardless of whether it is part of the disparate image that is responsible for the rivalry. In particular, the images of small test spots that are briefly projected onto the same retinal area are excluded. A rivalry scotoma can therefore be demonstrated using binocular perimetry, in which the visual field of one eye only is probed with conventional targets during binocular viewing with a dichoptic device. If the intensity of the stimulus within the rivalry scotoma is increased beyond a threshold level, it suddenly breaks through into visibility. Because the rivalry scotoma is present only under certain viewing conditions, it is described as facultative.
SENSORY FUSION
A retinal element is a small retinal patch that has an associated directional value. The fovea's directional value is defined subjectively as straight ahead; peripheral retinal elements possess directional values in other orientations. Corresponding retinal points are a pair of retinal elements, one in each eye, that have the same directional value. Comfortable single binocular vision occurs when objects in the binocular field (Fig. 5) stimulate corresponding retinal points and the higher cortical function (termed sensory fusion) occurs.
The locus in space that represents the intersection of all points in space that stimulate corresponding retinal points is termed the horopter (Fig. 6). Interestingly, sensory fusion still occurs if the object that projects upon a retinal element in one eye projects upon a range of elements that surrounds the corresponding retinal element in the second eye. The area in space that projects from this range of elements in the second eye that intersects with the projection from the retinal element in the first eye is termed the Panum fusional areas (Fig. 6). This Panum fusional area surrounds the horopter anteriorly and posteriorly; it permits fusion to take place when exact retinal correspondence does not occur. The binocularly perceived object imaged on noncorres-ponding retinal loci, but fused within the Panum fusional area, is perceived to have one subjective visual direction. The foveal Panum area is circular, of diameter about 14 min of arc; thus, an object projected upon the fovea of one eye may be displaced by this amount and the patient still maintains bifoveal vision. The size of the Panum fusional area increases toward the retinal periphery, but the ultimate size and shape depend upon the temporal and spatial frequency of the patient's alignment drift when fixing upon a stationary target (Fig. 6).
Objects in front of or behind the Panum fusional areas stimulate physiologic diplopia, which is not usually noted but may in turn stimulate fusional vergence eye movements. The horopter shape may be defined in a pair of perfectly spherical eyes that have refractive seats at the nodal points of each eye as the locus of points of zero vertical disparity relative to the fixation point. In a horizontal plane, the horopter, which includes the fovea, is the Vieth-Müller circle (Fig. 7). In a living animal visual system, the horopter is flatter (the Hering-Hellebrand horopter deviation). The vertical horopter tilts away from the observer, who stands on the horopter; the inclination is a function of fixation distance.



Fig. 2: Synoptophore—This instrument permits assessment of the angle alpha, the objective angle, abnormal retina correspondence, cyclophoria, hyperphoria and horizontal vertical vergences (Haag-Streit-UK)
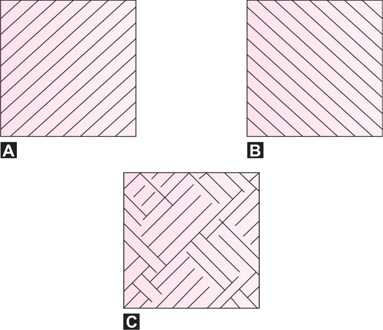
Figs 3A to C: Rivalry Pattern (A) Pattern seen by the left eye (B) Pattern seen by the right eye (C) Binocular impression. (Millodot: Dictionary of Optometry and Visual Science, 2009)

Fig. 4: Worth four-dot test consists of a black background and four illuminated disks: two green, one red and one white. Patient wears red/green glasses which makes each eye cancel the complimentary color (Gulden Ophthalmics)
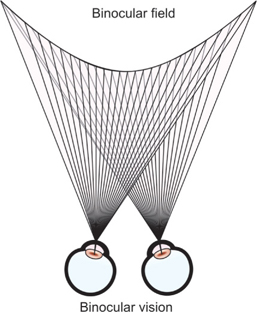
Fig. 5: Single binocular vision occurs when objects in the binocular field stimulate corresponding retinal points and sensory fusion occurs
STEREOPSIS
Stereopsis is the perception of the third dimension (relative nearness and farness of object points within Panum's area) obtained from fusible but disparate retinal images. It is possible to appreciate the relative location of objects using one eye (monocular cues).
Some monocular cues allow relative distance and depth to be judged. These monocular cues include:
- Relative size: Retinal image size allow us to judge distance based on our past and present experience and familiarity with similar objects. As the car drives away, the retinal image becomes smaller and smaller. We interpret this as the car getting further and further away. This is referred to as size constancy. A retinal image of a small car is also interpreted as a distant car.
- Interposition: Interposition cues occur when there is overlapping of objects. The overlapped object is considered further away.
- Linear perspective: When objects of known distance subtend a smaller and smaller angle, it is interpreted as being further away. Parallel lines converge with increasing distance such as roads, railway lines, electric wires, etc.
- Aerial perspective: Relative color of objects gives us some clues to their distance. Because of the scattering of blue light in the atmosphere, creating a “wall” of blue light, distance objects appear bluer. Thus, distant mountains appear blue. Contrast of objects also provides clues to their distance. When the scattering of light blurs the outlines of objects, the object is perceived as distant. Mountains are perceived to be closer when the atmosphere is clear.
- Light and shade: Highlights and shadows can provide information about an object's dimensions and depth. Because our visual system assumes that the light comes from above, a totally different perception is obtained if the image is viewed upside down.
- Monocular movement parallax: When our heads move from side to side, objects at different distances move at a different relative velocity. Closer objects move “against” the direction of head movement and farther objects move “with” the direction of head movement.
However, it is the lateral displacement of the eyes that provides two slightly different views of the same object (disparate images) and allows acute stereoscopic depth discrimination. The stereopsis determination hence is made on the basis of differences in interretinal distances of the fusible images within Panum's area: the greater interretinal image distances are projected by nearer object points, and the lesser interretinal image distances are projected by further object points.
Simultaneous binocular perception, fusion, and stereopsis are the three essential but distinctly different perceptual phenomena comprising single binocular vision. All three usually are capable of functioning simultaneously, although simultaneous perception is usually suppressed unless voluntarily recognized. There are some congenitally esotropic patients who, after their eyes have been straightened by surgery, only develop the simultaneous perception and fusion components of single binocular vision and never develop stereopsis. Binocular vision is a cortical function, and there are separate specialized cortical cells for each component. The cortical cells serving simultaneous perception, fusion, and stereopsis are conjectured to be completely individualized in their morphology, physiology, and distribution frequency throughout the cortex. They are indeed considered to be three separate neurophysiologic phenomena, sharing only the fact that they function only during the cortical processing of the images projected simultaneously on each retina.
There are two groups of clinical tests used to measure stereopsis. These are the contour stereotests and the random-dot stereotest.
Random-dot stereograms are pairs of images of random-dots which when viewed with the aid of a stereoscope, or with the eyes focused on a point behind the images, produce a sensation of depth, with objects appearing to be in front of or behind the actual images. The random-dot stereogram technique was first used by Dr Béla Julesz (1960) to eliminate monocular cues. As there are no contours, depth perception (stereopsis) can only be appreciated when binocular fusion occurs. Two process of stereopsis are used and these are local and global stereopsis. Local stereopsis exists to evaluate the two horizontally disparate stimuli. This process is sufficient for contour stereotests. Global stereopsis is required in random-dot stereogram when the evaluation and correlation of corresponding points and disparate points are needed over a large retinal area.
An example of a contour stereotest used in the clinic is the Titmus stereotest (Fig. 8). Vectograph cards dissociate the eyes optically. A vectograph consists of Polaroid material on which the two targets are imprinted so that each target is polarized at 90° with respect to the other. When the patient is provided with properly oriented Polaroid spectacles, each target is seen separately with the two eyes. This principle is used in the Titmus stereotest (Fig. 8).
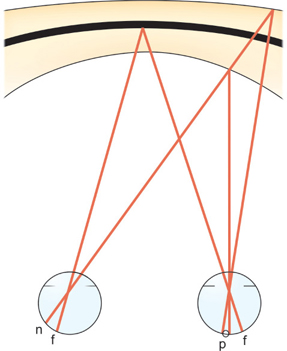
Fig. 6: Panum fusional area—The eyes are fixating “x” on the horopter “h”. Stimulation of point “n” in the left retina and of any point within Panum's area “p” of the right retina gives rise to a perception of singleness and stereopsis
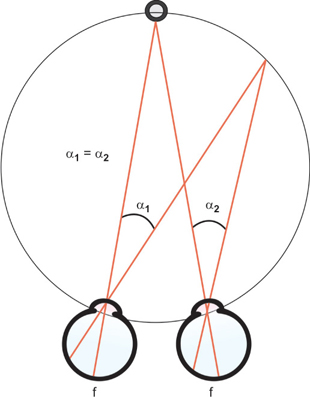
Fig. 7: Vieth-Müller Circle. This represents the theoretical location of points in space that will produce corresponding retinal points. Notice that the locations form a circle. If the eyes are assumed to be spherical with rotational centers at the nodal points, all points in space that have a zero disparity fall on this circle. Angle α1 = angle α2; thus, equal retinal distances map into equal angles in space in this idealized system

Fig. 8: Titmus stereotest. The standard for stereodepth perception testing. Evaluate both gross stereopsis (2500 to 1200 seconds of arc) and fine depth perception

Fig. 9: Frisby stereotest. This test consists of three test plates (6 mm, 3 mm, and 1.5 mm thick), which permit stereoacuity measurements in a range of 600 – 15 sec of arc. No special glasses are required. The hidden shape can only be detected if stereopsis is present. Suitable for a wide age range, even young pre-schoolers

Fig. 10: Randot stereotest with Polaroid glasses. Tests the ability to identify six geometric forms from random-dot backgrounds. Figures cannot be identified without glasses to discourage guessing. 500 to 20 seconds of arc
In this test a gross stereoscopic pattern representing a housefly is provided to orient the patient and to establish whether there is gross stereopsis (threshold: 3000 seconds of arc). In testing young children, one must ask questions the child will understand. For example, one may ask the child to take hold of the wings of the fly. If the child sees them stereoscopically, the child will reach above the plate. The Polaroid test also contains three rows of animals, one animal in each row imaged disparately (thresholds: 100, 200, and 400 seconds of arc, respectively). The child is asked which one of the animals stands out. The animal Figures contain a misleading clue. In each row one of the animals, correspondingly imaged in two eyes, is printed heavily black. A child without stereopsis will name this animal as the one that stands out. Last, the Titmus test (Fig. 8) contains nine sets of four circles arranged in the form of a lozenge. In this sequence the upper, lower, left, or right circle is disparately imaged at random with thresholds ranging from 800 to 40 seconds of arc. If the child has passed the other tests, he or she is now asked to “push down” the circle that stands out, beginning with the first set. When the child makes mistakes or finds no circle to push down, the limits of stereopsis are presumably reached.
Examples of random-dot stereotests used in the clinic are the Frisby stereotest (Fig. 9), the Randot stereotest (Fig. 10), the Random-dot E stereotest (Fig. 11) and the Lang stereotest (Fig. 12).
The Frisby stereotest has three transparent plates (Fig. 9). One of these is shown on each presentation to the subject. The target that the patient has to detect is printed on one side of each plate. This target is a cluster of randomly arranged arrowheads of differing sizes. On the other side of each plate is printed a background of similar texture. An observer with normal binocular stereovision can readily detect the target because it appears to stand out from the background (or to recede from the background, depending on which way round the plate is shown). Such an observer is essentially seeing the thickness of the plate by virtue of the texture elements printed on the two sides. An observer lacking normal binocular stereovision (or a normal observer viewing with one eye only) fails to be able to detect the target as it can be distinguished only on the basis of binocular disparity cues to depth.
Both the Randot (Fig. 10) and the Random-dot E stereotests (Fig. 11) use crossed polarized filters. Disparity is also constructed vectographically. The Randot stereotest uses modified animals and ring designs with random-dot backgrounds to eliminate monocular cues. The Lang stereotest uses a panographic technique (Fricke and Siderov, 1997) to present disparity; therefore, no filters are required. Patients are required to identify pictures on the Lang stereotest. The Lang II stereotest has a monocularly visible shape on it (Fig. 12).
All the tests provide a measure of stereoacuity by asking the patient to identify the correct target that has stereoscoptic depth (target with disparity). The working distance and interpupillary distance will need to be taken into consideration when calculating stereoacuity. Patients with disturbed binocular vision or different refractive error in one eye will perform poorly on depth discrimi-nation tests.
Patients with a large manifest deviation do not have useful stereopsis in casual seeing. Nevertheless, they can function quite well in space, making use of nonstereo-scopic clues to depth perception, especially if the strabismus is of early origin. They may have trouble with fast-moving objects, such as flying balls, and this experience may be frustrating to young children. However, when the strabismus is acquired later in life the loss of stereopsis is felt acutely and may present a real handicap. It appears as if stereopsis is useful in the comprehension of complex visual presentations and those requiring good hand-eye coordination.
Although the importance of stereopsis is often stressed, studies addressing the functional effects of stereoscopic deficits are sparse. It is always interesting and useful to determine whether a patient with strabismus has stereopsis or the potential for such.

Fig. 11: Random-dot E test set.(From Simons K, Reinecke RD: Amblyopia screening and stereopsis. In Symposium on strabismus: Transactions of the New Orleans Academy of Ophthalmology. St. Louis, Mosby–Year Book, 1978, p 15.)

Fig. 12: The Lang II stereotest. It measures disparities: Moon and Star are 200, car 400 and elephant 600 seconds of arc plus monocularly visible star. In this test, the random dots are smaller and less dense. This disparity is finer, namely 200 seconds of arc for the moon and the star, 400 for the car, and 600 for the elephant when viewed at a distance of 40 cm.